SEP
SNEST
DGEST
INSTITUTO
TECNOLOGICO DE CD ALTAMIRANO.
"REGULACIÓN DE LA EXPRESIÓN GENÉTICA".
LICENCIATURA EN
BIOLOGIA VI SEMESTRE.
MATERIA: BIOLOGIA
MOLECULAR.
ALUMNO: CANDIDO
TORRES SANTIBAÑEZ.
CON # DE CONTROL:
09930058.
PROFESOR: FRANCISCO
JAVIER PUCHE ACOSTA.
MAYO/ 2012.
OBJETIVO(S)
GENERAL(ES) DEL CURSO
Entenderá
las bases moleculares del control de la expresión génica y de la manipulación del
ADN mediante el empleo de las diferentes técnicas moleculares que le permitan
desarrollar proyectos de investigación básica y aplicada.
UNIDAD # 8 "Regulación de la expresión genética".
8.1 Niveles de regulación de la expresión genética.
8.2 Regulación de la transcripción en organismos
procarióticos.
8.4.1 Operon de Lactosa (Control positivo).
8.4.2 Operon de Triptofano (control negativo).
8.3 Regulación de la transcripción en organismos
eucarióticos.
La regulación genética comprende todos aquellos procesos que
afectan la acción de un gen a
nivel de traducción o transcripción, regulando sus productos funcionales.
Los organismos multicelulares complejos están compuestos de diferentes tejidos
cuyas características individuales dependen de las proteínas específicas
expresadas por sus tipos celulares. La diferenciación, el desarrollo y la
funcionalidad de los tejidos específicos dependen del conjunto de
proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas
en forma diferencial pueden funcionar como componentes estructurales de
las células, enzimas reguladoras del metabolismo, factores de
transcripción, receptores celulares, componentes intracelulares de
señalización, etc.
La expresión incorrecta de
tales proteínas, su expresión en lugares
equivocados, a destiempo,
o la producción en cantidades
anormales de proteínas específicas o de proteínas de función anómala subyace
a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la
expresión proteica en eucariontes contribuirá al conocimiento de las
bases moleculares de diversas patologías.
La metodología que se utiliza son los pasos a seguir para
la manifestación y desenlace de la unidad # 6 y con sus
correspondientes subtemas: realizando enlaces de acuerdo a lo manifestado
en clases de la unidad por el maestro y hacer conclusiones a lo entendido al
final de la unidad de todos los subtemas, publicando lo más interesante yde
cada tema visto en clase. y trabajos o ensayos obtenidos correspondiente a cada
clase.
EVIDENCIAS.
8.1 NIVELES DE REGULACIÓN DE LA EXPRESIÓN
GENÉTICA.
En las
células eucariotas, la capacidad de expresar proteínas biológicamente activas
resulta de diferentes niveles regulatorios.

La compactación de la cromatina afecta
la capacidad de unión de las enzimas y factores transcripcionales de genes
específicos. La cromatina se puede dividir en dos clases según su patrón de
tinción. La eucromatina se
tiñe suavemente y se corresponde con regiones del genoma que están
disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a
regiones del genoma que están densamente compactadas e inaccesibles para
el aparato transcripcional.
Se pueden distinguir dos clases de
heterocromatina: la
constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o
parte de ellos que son heterocromáticos en todas las células de una misma
especie, mientras que la facultativa implica zonas de cromosomas que se
pueden descompactar tornándose en eucromatina en algunas células de
un mismo organismo.
Como la heterocromatina no puede ser
transcripta, la expresión génica en los eucariontes se puede reprimir por
condensación de eucromatina en heterocromatina. Todavía no se conocen todos los
factores que modulan la descompactación de la cromatina. Ciertamente hay
proteínas que reconocen secuencias específicas del DNA y una vez unidas,
transmiten la señal de descondensación de cerca de 10000 pares de
bases correspondientes a un bucle de la cromatina.
Las
acetilaciones y desacetilaciones de histonas son modificaciones
covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un
ejemplo típico de este tipo de regulación ocurre en la acetilación
de coactivadores involucrados en las transcripciones genéticas
moduladas por las hormonas tiroideas. Las acetilaciones se producen en los
residuos de lisina de los extremos aminoterminales de las histonas, reduciendo
su carga positiva y por lo tanto su afinidad de unión al ADN cargado
negativamente. La desacetilación de las histonas, mediada por desacetilasas
provoca el efecto contrario (recompactación).
Secuencias características de
organización del DNA como los palíndromes así como la disposición
espacial del DNA Z han sido relacionados con
señalizaciones para el sitio de inicio de la transcripción.
Metilaciones
de residuos de desoxi citidina:
La metilación de los restos de citosina
en el ADN, especialmente en los sitios promotores, dificultan la transcripción.
Por ejemplo: los genes de globina están más metilados en células no productoras
de hemoglobina que en los eritroblastos. Las metilaciones se
producen en secuencias específicamente reconocidas ( 5’--- m CpG
---3’) que generalmente se agrupan en “islotes” ricos en GC, con frecuencia
dentro o cerca de regiones reguladoras de la transcripción.
La metilación puede inhibir la
transcripción de los genes al interferir en la capacidad de los factores
de transcripción para reconocer los sitios de unión al ADN o
alterando las conformaciones del ADN dificultando la polimerización de la
ARN polimerasa. Uno de los ejemplos más espectaculares de la metilación
ocurre durante el fenómeno de impresión genómica. Así, el conjunto
de cromosomas heredados del progenitor masculino no es funcionalmente
equivalente al conjunto de cromosomas heredados de la madre.Existen por
lo menos 100 genes sometidos a esta expresión diferencial. Las versiones
activas e inactivas de los genes difieren en sus patrones de metilación. Las
diferencias en los alelos se originan durante la gametogénesis.
Modificación
del número y de la estructura de los genes:
La eliminación
total o parcial de genes impide la formación de ARNm
y de la proteína correspondiente, los glóbulos rojos son un caso extremo donde
una vez sintetizadas las proteínas estructurales y funcionales, la eliminación
del núcleo en la etapa de eritroblasto ortocromático produce una célula incapaz
de sintetizar toda otra proteína de novo presentando un 90% del contenido
proteico total como hemoglobina.
Otro caso es la recombinación somática de la línea
germinal de los linfocitos B. En
este tipo particular de regulación de la expresión de genética, los genes
codificantes de las cadenas pesadas y livianas de la inmunoglobulinas sufren un
rearreglo independiente de la presencia del antígeno. Allí, se produce el corte
y empalme al azar de diversos fragmentos génicos de manera irreversible que dan
origen al variado repertorio de las inmunoglobulinas.
Por otra parte, la presencia de genes en tandem, implica la presencia de de múltiples copias
de un gen que aumentan la capacidad de producción de la proteína requerida en
grandes cantidades. Es el caso de los genes codificantes de histonas y
ARN 5S.
La regulación génica se puede regular
también en función de la disponibilidad del DNA incrementando el número de
copias de un gen accesible. Este mecanismo de regulación se conoce como amplificación génica. Una forma de amplificación es la repetición
sucesiva de la replicación de una secuencia específica del ADN. Este fenómeno
se observó en la amplificación de ciertos genes cuyos productos son
necesarios para el desarrollo de algunos insectos y anfibios. Como
ejemplo puede citarse el ARN ribosómico en la rana Xenopus laevis, donde los gnes que codifican los ARN r
5.8 S, 18S y 28 S se amplifican de 500 a 2 millones de copias.
Constituye uno de los modos más
importantes de regulación de la expresión proteica en eucariontes. En esta
categoría están incluídos los promotores, la presencia de secuencias
regulatorias potenciadoras (enhancers), y la interacción entre múltiples proteínas
activadoras o inhibidoras que actúan mediante su unión a secuencias específicas
de reconocimiento al ADN.
Las regulaciones
pueden ser de tipo CIS o TRANS.
Cuando el
elemento regulador transcripcional es parte de la cadena polinucleotídica donde
se localiza el gen a regular, se denomina regulador CIS. Evidentemente
se tratan de secuencias especiales del ADN (promotores y enhancers).
Cuando los elementos
regulatorios son de naturaleza y origen diferente a la secuencia genética
a controlar, la regulación es de tipo TRANS (aquí se incluyen a los factores de
transcripción generales, histoespecíficos y todas las proteínas
regulatorias con capacidad de unión al ADN).
Promotores
El paso inicial de la síntesis de los
tres tipos de ARN es la ubicación de las ARN polimerasas junto a una
secuencia del ADN a la altura del denominado promotor del gen a ser
transcripto. La ARN polimerasa I sintetiza los ARN r (excepto el 5S), la de
tipo II sintetiza los ARN m y algunos ARN sn involucrados en el proceso de
corte y empalme del transcripto primario (splicing), mientras que la ARN
polimerasa III sintetiza el ARN r 5S y los ARN t.El más complejo de los
procesos regulatorios es el que comprende a los genes de clase II o
codificantes de ARN m. Casi todos los genes codificantes de proteínas
contienen promotores basales de dos tipos y un número variable de
dominios regulatorios transcripcionales.
El promotor es
una secuencia que define el sitio de iniciación de la transcripción
del ARN.
Los promotores más frecuentes son los de
tipo CCAAT y TATA , denominados motivos o cajas por su alta conservación
evolutiva.
La caja TATA está localizada 20-30 pb
corriente arriba del sitio de inicio de la transcripción . Numerosas proteínas
identificadas como TF IIA,B,C, etc. (factores regulatorios de la ARN
polimerasa II) interaccionan con la caja TATA. El promotor CCAAT reside 50-130
pb corriente arriba del sitio de inicio transcripcional. La proteína denominada
C/EBP (CCAAT-box /enhancer/binding/protein) se
une a esta secuencia. Otra secuencia regulatoria incluye a la caja GC.
Si bien los promotores están
preferentemente localizados corriente arriba (5’) del inicio transcripcional,
algunos pueden ubicarse corriente abajo (3’) o bien ser de tipo intragénicos.
El número y tipo de eleemntos regulatorios varían según cada ARNm. La
naturaleza de los promotores y la combinatoria de las proteínas interactuantes
con ellos es uno de los principales mecanismos de regulación en los genes
de tipo inducibles.
Secuencias
regulatorias potenciadoras (enhancers):
Existen además secuencias del ADN que
pueden localizarse corriente arriba o abajo del gen a regular situadas a miles
de pares de bases con respecto al promotor. Las zonas de ADN sobre las cuales
se ejercen acciones activadoras son llamadas potenciadoras o
aumentadoras (enhancers en inglés). En términos generales una secuencia
regulatoria potenciadora o enhancer regula la frecuencia con la que se realiza
el proceso transcripcional.
Para un mismo gen pueden existir varias
secuencias regulatorias. También existen reguladores de acción opuesta
(silenciadores o amortiguadores de la transcripción). La explicación más simple
del fenómeno regulatorio a distancia es propone que la molécula de
ADN se dobla en asa para permitir la aproximación de estas zonas
alejadas de la doble hélice y ubica a la proteína activadora unida al
“enhancer".

Factores
transcripcionales:
A diferencia de las ARN polimerasas
bacterianas, las eucarióticas no hacen contacto directo con el ADN promotor
sino que son reclutadas hacia este por complejos de proteínas específicas
para cada tipo de ARN polimerasas. Estos complejos se han denominado SL1 para
la ARN polimerasa I, TFIID para la ARN pol, II y TF IIIB para la RNA pol III
respectivamente.
No hay duda alguna que los
factores transcripcionales de los genes codificantes de proteínas son de
crucial importancia en el control del crecimiento y la diferenciación de las
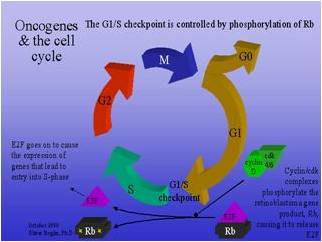
células. Muchos de ellos están codificados por protooncogenes, que al activarse
pueden alterar la tasa transcripcional o la calidad de las proteínas
fisiológicas desencadenando un proceso oncológico.
Se pueden distinguir los denominados factores de transcripción generales (componentes del complejo de
transcripción basal) y los factores de transcripción histoespecíficos (que se unen a regiones específicas
promotoras/reguladoras de los genes y regulan el grado de transcripción desde
el complejo de transcripción basal). Ambos tipos de proteína se unen a
secuencias en el surco mayor del ADN.
Para montar la transcripción basal
en un promotor dependiente de la ARN polimerasa II, se reclutan secuencialmente
factores de transcripción generales. El primer factor de transcripción general
que se une al promotor es el TFIID, que entra en contacto directo con la caja
TATA a través de la proteína fijadora de TATA (TBP). Una vez unido el
TFIIB los otros factores de transcripción generales (GTF, del inglés:
general transcription factors) se unen secuencialmente y por último se reclutan
la ARN polimerasas II y los factores asociados.
Este conjunto conformado por ADN,
factores de transcripción generales y ARN polimerasa II se denomina complejo de transcripción basal. Este
actúa como objeto de activación o represión transcripcional por la acción
de proteínas regulatorias específicas de tejido. Estos factores de
transcripción histoespecíficos se unen específicamente a otras
secuencias de ADN en las zonas de control de los genes intercalándose en el
surco mayor y comandan la interacción del complejo de transcripción basal
con el ADN, lo que provee una expresión génica regulada histoespecífica.
Además de su dominio de unión al ADN,
las proteínas reguladoras de la transcripción pueden tener dominios para
la interacción reversible, no covalente de una proteína con otra, denominados dominios de dimerización. La interacción de dos proteínas a través
de estos dominios puede ser un requisito para su unión al ADN. Por lo tanto, la
unión de factores de regulación es a menudo de tipo cooperativa Mientras muchos
factores diméricos poseen dos proteínas de la misma especie (homodímeros),
otros están formados por dos proteínas de diferente naturaleza (heterodímeros).
Es evidente que este tipo de proteínas
poseen además de dominios de dimerización, dominios de fijación al ADN y
dominios de activación transcripcional.
Constituye una zona proteica o dominio
compuesto por dos regiones de alfa hélice separadas por una de longitud
variable que forma un rulo o bucle entre ellas. Los dominios alfa helicoidales
son similares estructuralmente y necesarios para la interacción con secuencias
de la proteína que permiten una conformación simétrica respecto de un eje. Esta
clase de proteínas a menudo contiene una región rica en aminoácidos básicos
localizada en el extremo amino terminal que le permite su unión a secuencias
del ADN reconocidas específicamente.
Los dominios HLH son necesarios para
la homo y heterodimerización. Ejemplos para las proteínas de unión al ADN con
motivos HLH son: Myo D, c-Myc y Max. Varias proteínas que no contienen la zona
rica en aminoácidos básicos no se unen al ADN y actúan como represores de la
transcripción por esta causa. Así, estas proteínas HLH reprimen la
actividad de otras HLH por formación de heterodímeros y bloqueo de la unión al
ADN.

Este motivo consiste en espaciamientos
específicos conformados por residuos de cisteínas e histidinas que
permiten la unión de cationes Zn2+ a
la proteína, produciéndose un enlace coordinativo del metal en el centro de
ellos. Este fenómeno confiere al dominio aspecto de un dedo, por lo cual es
llamado comunmente “dedo de zinc”. Estos dominios pueden encajar en los surcos
mayores del ADN. El acople de estos factores regulatorios abarca la mitad de
una vuelta de la doble hélice del ADN. Los ejemplos más típicos son el
factor de transcripción de la ARN polimerasa II (TFIIIA) y las proteínas de la
superfamilia de receptores de las hormonas permisivas (esteroideas, tiroideas,
etc).

Es un motivo que se origina por la
distribución repetitiva de residuos de leucina espaciados por 7
aminoácidos en una distribución alfa helicoidal de la proteína. Estos residuos
de Leu terminan en una zona con residuos R que protruyen con respecto a
la zona helicoidal. Los grupos R se interdigitan con grupos R de otros
dominios de este tipo, estabilizando así la homo o heterodimerización. Los
dominios de cierre de lueucina están presentes en proteínas tales como: c-myc,
c-fos, c-jun y C/EBP.
La tasa de expresión de un gen es la
resultante de la interacción de varios de estos factores de transcripción
específicos que inducen o impiden la formación del aparato basal de
transcripción (compuesto por los factores de transcripción generales).
Los factores de transcripción específica
pueden de unión al ADN pueden activar o bloquear la transcripción si se fijan a
secuencias reguladoras consenso cercanas a la región del promotor. Si estas
proteínas se fijan a enhancers distantes de la región del promotor pueden aumentar
o disminuir la tasa transcripcional de un gen que se está transcribiendo.
La actividad de estos factores de
transcripción puede regularse a su vez a través de modificaciones covalentes
mediadas por cascadas de quinasas/fosfatasas y en algunos casos por
inducción/represión, dependiendo en todos los casos de señales externas. Esto
es muy importante para coordinar las funciones biológicas de la célula y su
interacción con el medio.

SITIOS ALTERNOS DE INICIO DE LA
TRANSCRIPCION: Un gen puede codificar para varias proteínas
al poseer dos exones 5’ diferentes ej: el gen de la amilasa y la
cadena ligera de miosina del ratón, la glucoquinasa y la alcohol deshidrogenasa
en la rata y la actina en la mosca de la fruta.
“PIEDRAS DEL CAMINO”: Una vez iniciada la transcripción, la
velocidad del movimiento de la RNA polimerasa puede reducirse e incluso
detenerse por periodos breves. Se asume que esto tiene que ver con el encuentro
de la polimerasa con determinadas secuencias del ADN llamadas “piedras del
camino” por su cualidad de detener la polimerasa. Se sabe que determinados
factores de transcripción favorecen el paso de la enzima por estos sitios y
otros provocan la liberación del aparato de transcripción de la plantilla,
determinando la finalización abrupta de la transcripción al interaccionar con
estos sitios.
Aunque el inicio de la transcripción es el
sitio primario de la regulación de la expresión de genes, la síntesis del
transcripto primario no es el único momento en que las células controlan la
producción adecuada de proteínas.
Puede ocurrir algo similar a la ocurrencia de
sitios alternativos de inicio de la transcripción pero en el extremo 3’. En
este caso hallándose sobre un mismo gen varios sitios susceptibles de ser
poliadenilados como es el caso del gen para la cadena pesada m de
inmunoglobulina. Dependiendo del sitio 3’ poliadenilado, la proteína resultante
puede anclarse a membrana o ser secretada, de acuerdo al estadío de desarrollo
en que se encuentra la célula.
Es un mecanismo muy difundido, que permite
que un solo gen pueda codificar para más de una proteína. En muchos de estos
casos se conoce más de una vía para procesar el transcripto primario obteniendo
proteínas estructural y funcionalmente diferentes o bien isoformas de una
proteína. Los cortes sobre el RNAhn deben producirse con absoluta precisión.
Puede ocurrir que uno más exones sean removidos (dando una proteína más corta),
o que uno o más intrones no sean removidos (dando una proteína más larga).
Se conoce
una familia de 6 proteínas llamadas ASF (factores de splicing alternativo)
responsables de reconocer y seleccionar los lugares para los cortes
alternativos, poseen la característica de contener dominios ricos en serina y
arginina por lo que se las llama también proteinas SR. El mecanismo por el cual
la célula selecciona los sitios no está claro. Los siguientes son algunos
ejemplos de genes cuya expresión puede regularse por splicing alternativo:
la a tropomiosina: esta se codifica a
partir de un mismo gen pero con una secuencia primaria diferente a partir de
ARNm diferentes específicos en 7 tejidos.
el gen de la tirosinhidroxilasa humana:
Esta enzima expresa 4 isoformas diferentes, es decir similar actividad
biológica con diferentes propiedades, en este caso se expresan todas en médula
suprarrenal y cerebro. El gen contiene 14 exones, al cortar y empalmar de
diferente manera, la isoforma 2 tiene 12 nucleótidos más que la isoforma
1, la isoforma 3 tiene 81 más y la isoforma 4 tiene 93 nucleótidos más que la
isoforma 1.
Los RNAm de las cadenas livianas de
inmunoglobulinas sufren corte y empalme alternativo, esto contribuye, como
veremos en inmunología, a ampliar el repertorio de anticuerpos, es decir a
reconocer más antígenos (elementos no propios) a los que pueda ser expuesto el
organismo. Así como permite mejorar la respuesta inmune a un antígeno en
particular.
Un mismo gen como el que codifica para
la hormona calcitonina –interviniente en la regulación de los niveles de Calcio
plasmático- al ser expresado en las células parafoliculares de la tiroides
cuando se expresa en las neuronas hipotalámicas produce una proteína llamada
CGRP (Producto relacionado al gen de calcitonina) que actúa como
neurotransmisor.
La fibronectina que cuando es
sintetizada por el fibroblasto y liberada a la matriz extracelular expresa dos
intrones que no son expresados cuando la proteína es sintetizada en los
hepatocitos y secretada al plasma.
Algunos genes codificantes para factores
de transcripción en células en etapas del desarrollo pueden ser ensamblados
alternativamente, la producción de una u otra variante determinará la vía de
diferenciación adoptada por la célula.
En la mayor parte de los casos, la diferencia
entre los productos de splicing alternativo sobre un mismo ARN difieren en
regiones claves que pueden afectar a la proteína en propiedades como: la
compartimentalización, el tipo de ligandos al que se unirá, la afinidad con que
lo hará y si es una enzima su actividad catalítica.
Si bien el splicing alternativo es la
forma mejor estudiada y más habitual de regulación de la expresión genética a
nivel de procesamiento del ARN, no es la única. La mutación puntual de
una base en el ARNnh o en el ARNm, puede alterar el producto. Esto no
constituye un error sino que es una herramienta utilizada por la célula para
lograr una determinada proteína. En el ser humano este es el caso de las
apoproteínas B100 y B48. Ambas son codificadas por el mismo gen y contienen en
el ARNnh los mismos exones e intrones. Cuando el gen se expresa en el
hepatocito, el ARNnh no sufre ningún tipo de modificación y el RNAm procesado
se traduce en ApoB100. En cambio, cuando el gen se expresa en el enterocito se
produce una mutación puntual que convierte C por U en el ARNm configurando un
codón de terminación que codifica para la ApoB48.
Los primeros estudios sobre el ARNnh
probaron que una fracción de éste es desdoblada en el núcleo por completo antes
de generar ARNm. Si bien aún los mecanismos no se conocen con certeza, los
estudios actuales sugieren que la célula puede controlar que un ARNnh sea o no
procesado. Este es el caso de la porción proteica de una
ribonucleoproteína, la U1A. Cuando ésta se expresa en cantidades altas se puede
ligar a su propio ARNnh e inhibir su poliadenilación de modo que el mensajero
resultante tiene baja estabilidad y disminuye así tu tasa de expresión.
ESTABILIDAD DEL ARNm:
A diferencia de los ARNm procariotas,
cuyas vidas medias rondan los 1-5 minutos, los ARNm eucariontes varían
ampliamente en su estabilidad. Por ejemplo: el ARNm de c-fos tiene una
vida media de entre 10 y 30’ en cambio el de las cadenas de globina es de más
de 24 hs.
La longitud de la cola de poli A está
directamente relacionada a la estabilidad citosólica del ARNm y esto se asocia
a la protección que ejerce esta secuencia al competir con el resto de la cadena
por la unión a nucleasas citosólicas. La cola de poli A se halla ligada a una
proteína llamada “Proteína de Enlace de Poli A” (PEPA) que protege a la
secuencia de la acción de nucleasas generales y la sensibiliza para nucleasas
específicas de poli A. A medida que estas nucleasas actúan la cola de poli A se
acorta hasta que ya no puede ligar a PEPA y el ARNm se vuelve susceptible a las
nucleasas generales y es rápidamente degradado.
Ciertos transcriptos inestables poseen
secuencias predominante pero no exclusivamente, en las regiones 3’ que
constituyen señales de rápida degradación para las nucleasas
citoplasmáticas. Estas secuencias se llaman “secuencias desestabilizadoras”,
constan de unos 50 nucleótidos y son ricas en A y U. Un grupo de nucleasas
citoplásmicas que se conocen como ribozimas y poseen un ARN pequeño en su sitio
activo pueden aparear con estas secuencias del extremo 3’ de manera específica,
acelerando la degradación del ARNm. Como ejemplo de estos transcriptos hallamos
a la mayoría de los ARNm para factores de crecimiento (caracterizados por una
vida media muy corta)
Ejemplos de
control de la estabilidad del ARNm en el citosol:
En el organismo humano los únicos
mensajeros no poliadenilados son los mensajeros de las histonas, que se
sintetizan constantemente a lo largo de todo el ciclo celular pero no se
transcribe debido a su gran inestabilidad (vida media de unos pocos minutos).
Cuando se progresa en G1/S lo que se incrementa es la tasa de transcripción de
los ARN de modo que al haber una gran cantidad disponible. se alcanza a
traducir. Una vez que la célula ha ingresado a la fase S la vida media del ARNm
de histonas se prolonga hasta 1 hora, posiblemente por la baja actividad de
nucleasas en este momento del ciclo celular.
La caseína, una proteína de la leche se
produce por la glándula mamaria en respuesta a prolactina. Aunque no se conoce
con certeza el mecanismo, bajo la acción de prolactina no se aumenta la tasa
transcripcional sino que se prolonga la estabilidad del mensajero.
La tubulina consta de dos subunidades (a y b)
que heterodimerizan. Cuando este dímero de tubulina alcanza determinada
cantidad en el citosol se une al extremo N terminal de los péptidos de
tubulina nacientes activando una nucleasa específica para los mensajeros de
tubulina, ejerciendo así un control de retroalimentación negativa sobre su
propia expresión.
La expresión de receptores de
transferrina también se maneja por estabilidad del mensajero. En este caso
particular, cuando las concentraciones citosólicas de hierro son bajas, el
mensajero liga en su extremo 3’ a la enzima aconitasa llamada también IRF (Factor
de respuesta al Hierro) Esto protege al ARNm de la acción de nucleasas. Cuando
la concentración citosólica de hierro aumenta, este compite con el mensajero
por la IRF se une al hierro y libera el extremo 3’ del mensajero del
receptor de transferrina. Este extremo 3’ posee 5 bucles con secuencias
desestabilizadoras que son rápidamente reconocidas por ribozymas. De este modo
se reduce la traducción de la proteína y con ello el ingreso de más hierro
–innecesario- a la célula.
Se descubrió también que los mensajeros
no se ubican al azar en el citosol En los fibroblastos humanos el 75% de los
ARNm poliadenilados se localizan en la intersección entre filamentos de actina
del armazón del citoesqueleto. Si bien no se conoce claramente la significación
de este hecho, en experimentos en la mosca de la fruta se comprobó que esta
disposición no azarosa tiene un papel muy importante en el proceso de la
fecundación.
Casi una tercera parte
del ARNm citoplásmico no está unido a ribosomas aún cuando más del 90% de los
ribosomas se encuentre en actividad. Se puede regular la velocidad con que los mensajeros son
traducidos y el número de veces que debe traducirse.
Existe un control general, global o
inespecífico de la traducción ejercido sobre los factores del complejo de
traducción. Por otro lado se conocen casos específicos o control particular de
la traducción de ciertos ARNm.
CONTROL GENERAL DE LA TRADUCCIÓN: Todos los ARNm celulares
tienen cap con la guanosina metilada y esa estructura aumenta la traducción.
Cuando por alguna razón de disposición espacial del extremo 5’ del ARN el cap
no queda disponible para su interacción el factor de iniciación Ife4E la
traducción disminuye. Puede controlarse también la actividad de los factores de
iniciación: IFe2B, IFe3, IFe4B e IFe4F. La actividad de estos factores se
controla por fosforilación. Esta fosforilación a su vez responde a diversos
estímulos que pueden ser fisiológicos (factores activadores de cascadas
de quinasa) o no ( desnutrición severa, hiperosmolaridad, infección viral
o shock térmico)
El IFe2B parece ser entre ellos el más
importante desde el punto de vista del control de la traducción. En el caso
particular del IFe2B es la subunidad a la susceptible de ser fosforilada por
una proteinquinasa independiente de AMPc que responde a señales de
estrés y se inactiva evitando la formación del complejo 40S.
El IFe4F puede ser fosforilado por
Insulina y Cascadas de quinasa activadas por mitógeno en este caso la
fosforilación activa al factor promoviendo un aumento en la traducción. Existen
2 proteínas (4E-BP1 y 4E-BP2) que se ligan al IFe4F inactivándola. La insulina
y los factores de crecimiento pueden fosforilarlas liberando al IFe4F y dejando
disponibles los sitios de fosforilación para potenciar la estimulación de la
actividad del factor y con ello la síntesis de proteínas.
CONTROL PARTICULAR DE LA TRADUCCIÓN : Depende de sustancias reguladoras que
modifican la configuración de un tramo de nucleótidos no traducibles
localizados en el extremo 5’ entre el cap y el codón de iniciación.
Un ejemplo de este tipo de regulación lo
proporciona la traducción del mensajero de la Ferritina que se une al hierro
para su depósito intracelular. Cuando las concentraciones citosólicas de hierro
aumentan la síntesis de ferritina se produce a altas velocidades, el hierro se
liga a la aconitasa o IRF. En cambio, cuando estas concentraciones disminuyen
la IRF queda libre y se liga al ARNm de la ferritina formando un bucle en el
extremo 5’ que impide el ligado de IFe4F y con ello el ensamblaje del aparato
de traducción, bloqueando de esta manera la producción innecesaria de proteína.
Otro ejemplo podemos verlo en los
reticulocitos en que la síntesis del hem está controlada por el propio grupo
hem. Cuando hay suficiente hem, este activa a la proteinquinasa de IFe2B
regulada por hem que inactiva al factor por fosforilación.
En los últimos años se han descubierto
nuevos procesos en cuanto a control de la traducción que operan en algunas
células en particular. Ellos son
CAMBIO DEL MARCO DE LA TRADUCCIÓN: es
decir, el ribosoma cambia la pauta de lectura en algún punto de su movimiento a
lo largo del ARNm desplazándose un nucleótido hacia atrás o hacia delante,
pudiendo, un mismo mensajero dar origen a dos proteínas.
SALTO DEL CODÓN DE TERMINACIÓN: el
ribosoma pasa por alto el codón de terminación y continúa leyendo la secuencia
de nucleótidos.
PASO POR ALTO DE LA TRADUCCIÓN: el
ribosoma “salta” una secuencia de nucleótidos en el mensajero de modo que queda
un mensajero más largo que la proteína porque resta una porción interna del
mensaje sin traducir, pudiendo nuevamente un mismo mensajero dar origen a dos
proteínas
EMPALME DE PROTEÍNAS: Al modo del
splicing se conocen casos en que un segmento específico del polipéptido se
secciona y los dos bordes se unen en forma covalente.
El péptido naciente
del ribosoma puede sufrir diversas modificaciones acorde con su función
biológica posterior incluyendo, remoción del extremo amino terminal, agregado
se secuencias señal para exportación, acilaciones, metilaciones, sulfataciones,
glicosilaciones, prenilaciones, modificaciones dependientes de vitamina C,
carboxilaciones dependientes de vitamina K –como en el caso de los factores de
coagulación- o bien clivajes post-traduccionales para tornarse activas
(lo cual es un modo de controla su actividad como en el caso de algunas enzimas
y hormonas peptídicas).
Todas estas
modificaciones pueden ser controladas por señales internas (pH intracelular,
actividad de chaperonas moleculares) o externas (cascadas de quinasa que
controlan fosforilaciones) Cualquier problema en estas modificaciones
post-traduccionales puede desencadenar alteraciones en la fisiología de la
célula. La actividad post-traduccional se controla a través de la regulación de
la actividad de las enzimas intervinientes en los procesos citados.
Cualquiera sea la
estructura y función de la proteína es indispensable un correcto plegamiento
post-traduccional de los péptidos ya que la formación de estructuras 2º,
suprasecundarias y 3º, así como las oligomerizaciones serán las responsables de
que la proteína sea funcional.
Algunos mensajeros codifican para
poliproteínas. Éstas son productos transitorios que se escinden en varias
proteínas, cada una con estructura y función diferente. Un ejemplo de
poliproteína es el producto del gen POMC (Proopiomelanocortina) que puede escindirse
en modo diferencial según sea que esté en las células adrenocorticotropas del
lóbulo anterior de la hipófisis –generando ACTH y b lipotrofina-
o en las células de la pars intermedia donde genera bendorfina, glipotrofina y hormona estimulante de
Melanocitos en sus dos formas aMSH
y bMSH.
Las células pueden controlar el tiempo
de supervivencia de las proteínas una vez que han sido sintetizadas y
modificadas post-traduccionalmente. Aunque no se trata directamente de la
regulación de la expresión de genes, es una extensión del tema.
Los mecanismos que controlan la
estabilidad proteica no están bien comprendidos, aunque se sabe que en las
proteínas que en el extremo amino terminal son ricos en metionina, serina,
alanina, treonina, valina y glicina son estables por más de 20hs en cambio los
que son ricos en fenilalanina, ácido aspártico, lisina y arginina tienen una
vida menor a 5’. Esto se llama “Regla del Extremo N”. Las últimas
secuencias mencionadas se llamas “Secuencias PEST” – por la nomenclatura
internacional de aminoácidos- y son altamente sensibles al reconocimiento por
ubiquitina y con ello su degradación en el proteasoma.
TERMINOLOGÍA
|
Unidad de transcripción
|
Los
genes/cistrones/ORF que se transcriben bajo en control de un mismo promotor
|
Promotor
|
Secuencia
reconocida específicamente por la RNA polimerasa para iniciar la
transcripción de una unidad de transcripción.
|
Cistrón
|
Cada una
de las partes de un transcrito que darán una biomolécula funcional (proteína
o RNA). En el caso de las proteínas, coincide con un marco abierto de lectura
(ORF).
|
Terminador
|
Conjunto
de secuencias que marcan el fin de la transcripción de una unidad de
transcripción.
|
Operón
|
La
unidad de transcripción (promotor, cistrones y terminador) junto con las
secuencias/genes adicinales que sirven para regularlo.
|
Polaridad
|
Es el
efecto de una mutación en un gen sobre la expresión de otros cistrones
distales dentro del mismo operón
|
Proteína reguladora
|
Aquella
que controlará la expresión de un operón. Pueden ser activadoras o represoras.
|
Operador
|
Secuencia
del promotor que es reconocida por una proteína reguladora.
|
Efector
|
Biomolécula
no proteica que controla la activación o inactivación de la proteína
reguladora. Las hay inhibidoras y (co-)represoras.
|
Regulón
|
Conjunto
de genes/operones que responden al unísono por la acción de un regulador. Por
ejemplo los regulones de choque térmico, represión catabólica, o respuesta
SOS.
|
Cambios
en la estructura del DNA
-Metilación
La metilación provoca un cambio de estructura en el apareamiento
entre las bases nitrogenadas que puede alterar su reconocimiento por algunas
proteínas. El más conocido es el de la metilasa dam que reconoce la secuencia
GATC y metila la A.
La mayor parte de los genes cuya exprexión se ve reprimida por la
metilación son genes cuya expresión sólo se necesita durante la replicación
(único momento en el que una cadena del DNA está transitoriamente
hemimetilado), permaneciendo reprimidos el resto del ciclo celular.
Los genes mioC y dnaA de E. coli implicados en el inicio de la
replicación del DNA son de los pocos que se activan con la metilación, puesto
que si el DNA está totalmente metilado, es señal de que se pueden expresar para
iniciar otro ciclo replicativo.
-Superenrollamiento
Para mantener una situación homeostática en la célula en relación
al número de superenrollamientos es necesario mantener con una regulación contraria
los genes topA (codifica la topoisomerasa I) y gyrA y gyrB (determinan las dos
subunidades de la DNA-topoisomerasa II). No se conoce el mecanismo molecular
que controla esta regulación. Sí se sabe que mutantes en las topoisomerasas
disminuyen la tasa general de transcripción.
Superenrollamiento
Para mantener una situación homeostática en la célula en relación
al número de superenrollamientos es necesario mantener con una regulación
contraria los genes topA (codifica la topoisomerasa I) y gyrA y gyrB
(determinan las dos subunidades de la DNA-topoisomerasa II). No se conoce el
mecanismo molecular que controla esta regulación. Sí se sabe que mutantes en
las topoisomerasas disminuyen la tasa general de transcripción.
Cambios
en la interacción entre el DNA y la RNA-polimerasa
Lo provocan aquellos cambios que, sin alterar ni la estructura del
DNA ni la de la RNA-polimerasa, sí que afectan la interacción entre ambas. Es
necesaria la comparecencia de una tercera molécula, habitualmente una proteína
(aunque a veces puede ser RNA). Tres mecanismos pueden alterar esta
interacción:
• Modificación de la
interacción entre RNA polimerasa y el promotor debido a la existencia de una
proteína reguladora y un efector
• Secuencias
reguladoras a distancia
• Modificación de la terminación:
antiterminación.
8.2.1
OPERON DE LACTOSA (CONTROL POSITIVO).
Control positivo: Se dice que un sistema está bajo control positivo cuando el producto del gen regulador activa la expresión de los genes, actúa como un activador.
ELEMENTOS
DEL OPERÓN
Jacob,
Monod y colaboradores analizaron el sistema de la lactosa en E. coli, de
manera que los resultados de sus estudios permitieron establecer el modelo
genético del Operón que permite comprender como tiene lugar la
regulación de la expresión génica en bacterias. Jacob y Monod recibieron en
1965 el Premio Nobel pos estas investigaciones.
FOTO DE ARRIBA ES: FLACOIS JACOB Y EL DE ABAJO ES: Jacques Monod
Un Operón es grupo de genes estructurales cuya expresión está regulada por los mismos elementos de control (promotor y operador) y genes reguladores.
Los
principales elementos que constituyen un operón son los siguientes:
v Los genes estructurales: llevan información para polipéptidos. Se trata de los genes cuya
expresión está regulada. Los operones bacterianos suelen contener varios
genes estructurales, son poligénicos o policistrónicos. Hay algunos operones
bacterianos que tienen un solo gene estructural. Los operones eucarióticos
suelen contener un sólo gen estructural siendo monocistrónicos.
|
v El promotor (P): se trata
de un elemento de control que es una región del ADN con una secuencia que es
reconocida por la ARN polimerasa para comenzar la transcripción. Se encuentra
inmediatamente antes de los genes estructurales. Abreviadamente se le designa
por la letra P.
|
v El operador (O): se trata
de otro elemento de control que es una región del ADN con una secuencia que
es reconocida por la proteína reguladora. El operador se sitúa entre la
región promotora y los genes estructurales. Abreviadamente se le designa por
la letra O.
|
v El gen regulador (i): secuencia de ADN que codifica para la proteína reguladora que reconoce
la secuencia de la región del operador. El gen regulador está cerca de los
genes estructurales del operón pero no está inmediatamente al lado.
Abreviadamente se le denomina gen i.
|
v Proteína reguladora: proteína
codificada por el gen regulador. Está proteína se une a la región del
operador.
|
v Inductor: sustrato
o compuesto cuya presencia induce la expresión de los genes.
|
| Elementos que intervienen en la regulación de la expresión
génica en bacterias. Elementos del Operón. |
Elementos de control
|
Promotor
|
Operador
|
Moléculas difusibles
|
Proteínas reguladoras
|
Efectores
|
Inductores
|
Genes Estructurales
|
Codifican para polipéptidos
|
Gen regulador
|
Codifica para proteína reguladora
|
OPERÓN LACTOSA: CONTROL POSITIVO
Sin embargo,
en presencia del inductor (la lactosa), este se une a la proteína reguladora
que cambia su conformación y se suelta de la región operadora dejando acceso
libre a la ARN-polimerasa para que se una a la región promotora y se
transcriban los genes estructurales. Por consiguiente, la presencia del
inductor hace que se expresen los genes estructurales del operón, necesarios para
metabolizar la lactosa.
Operón lactosa en presencia de lactosa
|
También es conveniente recordar que los tres genes estructurales del operón lactosa se transcriben juntos en un mismo ARNm, es decir que los ARN mensajeros de bacterias suelen ser policistrónicos, poligénicos o multigénicos. Sin embargo, en eucariontes los mensajreos suelen sen monocistrónicos o monogénicos, es decir, corresponden a la transcripción de un solo gen estructural.
8.4.2 OPERON DE TRIPTOFANO.
El operón triptófano (operón trp) es un sistema de tipo
represible, ya que el aminoácido triptófano (Correpresor) impide la expresión
de los genes necesarios para su propia síntesis cuando hay niveles elevados de
triptófano. Sin embargo, en ausencia de triptófano o a niveles muy bajos se
transcriben los genes del operón trp. Los elementos del operón trp son en
esencia semejantes a los del operón lactosa:
·
Genes
estructurales:
existen cinco genes estructurales en el siguiente orden
trpE-trpD-trpC-trpB-trpA.
|
·
Elementos de control: promotor (P) y operador (O). El promotor y el operador están al lado de
los genes estructurales y en el siguiente orden: P O trpE-trpD-trpC-trpB-trpA.
Curiosamente, las enzimas codificadas por estos cinco genes estructurales
actúan en la ruta metabólica de síntesis del triptófano en el mismo
|
·
Gen regulador (trpR): codifica para la proteína reguladora. Este gen se encuentra en otra
región del cromosoma bacteriano aunque no muy lejos del operón.
|
·
Correpresor: triptófano
|
En el siguiente esquema se indican
los elementos del Operón Triptófano:
Elementos del Operón Triptófano
|
Los genes
estructurales del operón triptófano se encuentran en el mismo orden que actúan
las productos codificados por ellos en la ruta biosintética del triptófano (ver
el siguiente esquema).
Orden de los
genes estructurales del operón triptófano y ruta de síntesis del triptófano
Operón
triptófano: en ausencia de triptófano
En ausencia
de triptófano, o cuando hay muy poco, la proteína reguladora producto del gen trpR no es capaz de unirse al
operador de forma que la ARN-polimerasa puede unirse a la región promtora y se
transcriben los genes del operón triptófano.
Operón
triptófano: en presencia de triptófano. Por tanto, la diferencia esencial entre
el operón lac (inducible) y el operón trp (represible), es que en este último
el represor del triptófano solamente es capaz de unirse al operador cuando
previamente está unido al trp.
Al estudiar más profundamente el operón trp se
encontró que además del mecanismo de regulación represor-operador existía otro
mecanismo de regulación que se denominó regulación por atenuación.