INSTITUTO TECNOLOGICO DE CD ALTAMIRANO

UNIDAD # 5 "Reparación del material genético".
QUE PRESENTA:
ALUMNO: Candido torres santibañez.
CON # DE CONTROL: 09930058.
PROFESOR: Francisco javier puche acosta.
CD ALTAMIRANO, GUERRERO. MEXICO. A 23/ 04/2012
UNIDAD # 5. "REPARACIÓN DEL MATERIAL GENÉTICO".
5.1 Clasificación de los tipos de lesión al ADN
5.1.1 Lesiones espontáneas
5.1.2 Lesiones inducidas
5.1.2.1 Fijación de la lesión
(mutación)
5.1.2.2 Agente mutagénicos
5.1.3 Físicos
5.1.4 Químicos.
5.2 Sistemas de reparación.OBJETIVOGENERAL DEL CURSO
Entender las bases moleculares del control de la expresión génica y de la
manipulación del ADN mediante el empleo de las diferentes técnicas moleculares
que le permitan desarrollar proyectos de investigación básica y aplicada.OBJETIVO EDUCACIONAL DE LA UNIDAD # 5
Definir la mutación,su papel en la genética molecular y los distintos tipos que
se conocen.
Relacionar los distintos mutágenos con los defectos en la organización del
genoma.
Conocer los mecanismos de reparación molecular del material genética
de los seres vivos con el fin de entender La estabilidad y la
variabilidad genéticaINTRODUCCIÓN.
En
esta dicha unidad que es la # 5.se comprendera las distintas modalidades en el que el
material genético repara los daños. Los
daños en el ADN son reparados para mantener la integridad de la información
genética, la importancia biológica de la reparación del ADN es evidente al encontrar
múltiples mecanismos de reparación.
El
material genético está constantemente
expuesto a agentes que le causan daño (los agentes físicos)
tales como la radiación y (los agentes químicos). Por
otra parte una de las fuentes de
variabilidad genética que han hecho posible la evolución es la mutación o
cualquier cambio heredable en la secuencia de nucleótidos del material genético (ADN) de
un organismo. Las mutaciones suponen la alteración del genotipo, o constitución
genética del individuo, y en ocasiones también del fenotipo que son las
características externas del individuo.
Una de las fuentes de variabilidad genética que han hecho posible la evolución es la mutación o cualquier cambio heredable en la secuencia de nucleótidos del material genético (ADN) de un organismo.
Las mutaciones suponen la alteración del genotipo, o constitución genética del individuo, y en ocasiones también del fenotipo que son las características externas del individuo.
Las mutaciones ocurren al azar, esto se descubrió en un experimento en el que se hicieron 10 cultivos de 10 (8) células cada uno.
A cada cultivo se le añadió el fago T1, las células eran de E. coli.
Si las mutaciones no ocurren al azar cabría esperar que el número de colonias resistentes fuera más o menos igual en cada cultivo, si la mutación es al azar se espera gran variabilidad en el número de colonias resistentes en cada tubo.
Así, se comprobó, que la mutación era al azar.
Las mutaciones pueden afectar a uno (puntuales), unos pocos (pseudopuntuales) o a un gran número de nucleótidos de una secuencia de ADN (cromosómicas).
Puntuales y pseudopuntuales
* cambios de base
- transiciones: Purina por purina y pirimidina por pirimidina
- transversiones: Purina por pirimidina y pirimidina por purina
- deleción
- inserción
- deleciones
- duplicaciones
- inversiones
- translocaciones
MUTACIONES ESPONTÁNEAS
Errores en la replicación del DNA
Durante la síntesis del DNA puede producirse un error en la replicación porque se forme un emparejamiento ilegítimo de nucleótidos como A-C que da lugar a la sustitución de una base por otra.
Cada una de las bases aparece en el DNA en una de varias formas llamadas tautómeros que son isómeros que se diferencian en las posiciones de sus átomos y en los puentes que se forman entre ellos. Esas formas están en equilibrio.
La forma ceto es la que se encuentra normalmente en el DNA mientras que las formas imino o enol son menos frecuentes.
La capacidad del tautómero menos frecuente de una base de emparejarse erróneamente y producir mutaciones durante la replicación del DNA fue puesta de manifiesto por primera vez por Watson y Crick. A estos emparejamientos erróneos se les llama cambios tautoméricos.
También pueden ocurrir emparejamientos erróneos cuando una de las bases se ioniza, esto sucede con más frecuencia que los cambios tautoméricos.
Transiciones
Todos los emparejamientos erróneos anteriores producen mutaciones por transición, en las que una purina es sustituida por otra purina y una pirimidina es sustituida por otra pirimidina.Transversiones
No pueden realizarse por emparejamientos erróneos como los debidos a cambios tautoméricos.Pero sí pueden realizarse si una base sufre un cambio tautomérico mientras que la otra base rota sobre su enlace glucosídico y quedan enfrentadas sus cargas.
Desaminación
Es una de las más frecuentes debido a la inestabilidad química, afectando gravemente a la replicación del ADN provocando transiciones.En este caso la base se modifica antes de la replicación debido a los radicales que provoca el metabolismo.
La desaminación de citosina produce uracilo, así los resíduos de uracilo que no sean reparados se emparejarán con adenina durante la replicación produciendo la conversión de un par GC en uno AT, se produce una transición.
Cambios de fase
Estas mutaciones pueden ser inserciones o deleciones.Las inserciones se producen por un deslizamiento o "resbalón" de la cadena sintetizada con lo que se forma un lazo de varios pares de bases.
En la siguiente ronda de replicación se añadirán tantas bases como comprenda el lazo ya que cuando se produce el "resbalón" sigue replicándose por donde se quedó antes del "resbalón".
Las deleciones se producen por un deslizamiento o "resbalón" de la cadena molde, como las que hay que copiar no se pueden no se añaden a la caden hija.
Despurinización
El ADN pierde de alguna manera alguna de sus bases y si hay un hueco la reparación introduce una base.La frecuencia de las mutaciones espontáneas es generalmente baja.
EFECTOS DE LOS CAMBIOS
Se expresan cuando el gen pasa a su proteína correspondiente.Los efectos de los cambios pueden ser:
- Cambios de sentido: se cambia un aminoácido por otro
- Sin sentido: la mutación se produce porque se transforma en un codón de terminación.
- Desfases: si hay una deleción de la base, la pauta de lectura cambia y se produce un gran cambio en la proteína y es muy grave.
- Mutaciones silenciosas: son mutaciones sin efecto: UUU (Phe)---> UUC (Phe)
- El aminoácido que cambia es muy parecido y la proteína sigue funcionando.
MUTACIONES INDUCIDAS
Existen puntos de un gen donde la mutación es más frecuente se llaman PUNTOS CALIENTES. Al genotipo silvestre o salvaje se le utiliza como patrón y en el que se produce la variación se le llama mutante.Una estirpe mutante puede cambiar a otra y luego volver a la inicial, a esto se le llama regresión. Los mutantes se inducen con mutágenos que son de varios tipos y cada uno induce una mutación distinta, aunque suele ser al azar.
Los mutágenos son de varios tipos:
Mutágenos Químicos
Análogos de bases:
Algunos compuestos químicos son suficientemente parecidos a las bases nitrogenadas normales del DNA para, ocasionalmente, incorporarse a éste en lugar de las bases normales, tales compuestos se llaman análogos de bases.
Una vez en su sitio tienen propiedades de emparejamiento distintas de aquellas a las que han sustituido, de este modo, causan mutaciones al provocar que, durante la replicación, se inserten frente a ellas nucleótidos incorrectos.
El análogo de base original sólo están en una cadena sencilla pero puede provocar el cambio de un par de nucleótidos que se replica en todas las copias de ADN descendientes de la cadena original. Ejemplos son: 5-bromurouracilo, 2-aminopurina.
Modificadores de bases:
- ácido nitroso: provoca una desaminación que modifica las bases C-->U, G--->X, con lo que se produce un apareamiento erróneo.
- Hidroxilamina: provoca una transición de G-->A y se da principalmente en bacterias.
- Agentes alquilantes: introducen grupos alquilo a las cuatro bases en muchas posiciones, produciendo transiciones, etilmetanosulfonato y la nitrosoguanidina.
- Agentes intercalantes: son moléculas planas que imitan pares de bases y son capaces deddeslizarse entre las bases nitrogenadas apiladas en el núcleo de la doble hélice, mediante un proceso de intercalación. En esta posición el agente puede producir deleciones o deleciones de un par de nucleótidos. Algunos agentes intercalantes son: proflavina, naranja de acridina y ICRs.
Pérdida del emparejamiento específico:
Un gran número de mutágenos dañan una o más bases, haciendo imposible el posterior emparejamiento específico. El resultado es un bloqueo en la repliación, puesto que la síntesis del DNA no sigue más allá de una base que no puede especificar una complementaria mediante puentes de hidrógeno. Este fallo es replicado por el mecanismo SOS.Radiaciones
UV que producen dímeros de timina, rayos X y las radiaciones gamma que rompen el DNA.MUTACIÓN Y REPARACIÓN DEL ADN. TEST DE AMES
Es un test para detectar la carcinogenicidad. Utiliza dos mutaciones de auxotrofía para histidina que revierten por diferentes mecanismos moleculares. Llevan una mutación que inactiva el sistema de reparación por escisión, y otra que elimina la cubierta protectora.SUPRESIÓN Y REVERSIÓN
Se explica mediante un ejemplo. Si tenemos una mutante de E. coli que no crece en un medio sin histidina, es his- y es auxótrofo.El silvestre se llama his+.
Sometemos el mutante a mutágenos y puede pasar a his+, y es una reversión. En este mutante puede ocurrir una reversión verdadera o una reversión equivalente.
Se produce una supresión cuando la segunda mutación se produce en otro sitio pero sí se convierte en his+.
Es una complementación intergénica.
La supresión intergénica consiste en una mutación en otro gen distinto de donde ocurrió la primera.
La supresión intragénica consiste en una mutación supresora en el mismo gen que ocurrió la supresión inicial.
La complementación intragénica se produce sobre todo en proteínas polímero.
MECANISMOS DE REPLICACIÓN
Reparación directa
Son sistemas que eliminan directamente el daño del UV en el DNA, como es el caso de los dímeros de timina. La luz visible activa la fotoliasa que rompe los dímeros de timina.
Otro ejemplo son las alquiltransferasas y su actividad consiste en eliminar los grupos alquilo, también se repara la despurinización gracias a las glicosidasas del ADN.
Dependiente de replicación
Todas las células contienen endonucleasas que atacan los sitios que quedan tras la pérdida espontánea de resíduos de purina o pirimidina. Por comodidad, los sitios sin purina o sin pirimidina se denominan sitios AP.Las endonucleasas AP son vitales para la célula porque, como se apuntó con anterioridad, la despurinización espontánea es un hecho relativamente frecuente.
Estas enzimas introducen hendiduras en la cadena mediante la rotura de enlaces fosfodiésteres en los sitios AP. Esto promueve un proceso de reparación por escisión medidado por otras tres enzimas: una exonucleasa, la polimerasa de DNA I y la ligasa de DNA.
Escisión
Esta vía de reparación está determinada por tres genes denominados uvrA, uvrB y uvrC. Este sistema reconoce cualquier lesión que cree una distorsión importante en la doble hélice de DNA. Una endonucleasa denominada nucleasa uvrABC realiza una incisión alejada varios pares de bases a cualquier lado de la base dañada, eliminándose a continuación un fragmente de DNA de cadena sencilla. El pequeño hueco se rellena entonces mediante síntesis de reparación y queda sellado por la ligassa de DNA.Sistema GO
Dos glucosilasas actúan conjuntamente para eliminar las mutaciones causadas por las lesiones que produce en el DNA el 8-oxodG. Las glucosilasas junto al producto del gen mutT forman el sistema GO.Cuando se originan lesiones GO en el DNA, por daño oxidativo espontáneo, una glucosilasa cifrada en el gen mutM elimina la lesión.
Aún así persisten algunas lesiones GO que emparejan erróneamente con adenina. Una segunda glucosilasa producto del gen mutY elimina la adenina de este emparejamiento erróneo específico, llevando al restablecimiento de la citosina correcta por síntesis de reparación.
Sistema SOS
En E. coli depende de los genes recA, umuC y umuD. Cuando se encuentra un tramo sin cifrar actúa el sistema SOS.Se activa la proteína recA que induce la presencia de las proteínas SulA y SulB que interaccionan con la DNA pol III. Ésta hace que pierda afinidad y prosiga la síntesis de ADN y dejando el hueco y sin que la célula muera.
Reparación postreplicativa
Algunas vías de reparación reconocen errores incluso después de que haya tenido lugar la replicación. Uno de estos sistemas, denominado sistema de reparación de emparejamientos erróneos.Para averiguar cual de las dos bases es la errónea debe diferenciar entre la cadena progenitora y la cadena hija.
Lo diferencia porque la enzima metiladora metilasa de la adenina tarda varios minutos en metilar la cadena hija.
Las proteínas mut S y mut L interaccionan con el sitio mal emparejado y una proteína mutH rompe la cadena recién sintetizada. Alrededor del emparejamiento erróneo, las cadenas de DNA se separan con ayuda de una proteína denominada MutU y se estabilizan con SSB. Y las polimerasas copian el segmento de DNA.
METODOLOGIA.
5.1 Clasificación de
los tipos de lesión al ADN
Restricción: mediante actividades metilasa y endonucleasa se protege el DNA propio
del foráneo.
Recombinación: se redistribuyen o reorganizan dos moléculas de DNA
Reparación: corrige aquellos errores introducidos en la secuencia del DNA tras la replicación
Transposición: es un tipo especial de recombinación con el que se consigue cambiar de posición un DNA, o sea, reorganizarlo, y en otras lo que hace es amplificarlo.
Recombinación: se redistribuyen o reorganizan dos moléculas de DNA
Reparación: corrige aquellos errores introducidos en la secuencia del DNA tras la replicación
Transposición: es un tipo especial de recombinación con el que se consigue cambiar de posición un DNA, o sea, reorganizarlo, y en otras lo que hace es amplificarlo.

5.1.1 Lesiones
espontáneas
Se consideran alteraciones o
lesiones todos aquellos procesos en los que el DNA es sustrato de la reacción y
no molde. Además, al final del proceso el DNA resultante es distinto al DNA inicial.
Lesiones espontáneas son entonces los errores que aparecen
de manera natural en los procesos de copia del ADN o en el trascurso de la vida
celular de un organismo.
5.1.2 Lesiones
inducidas
Varias actuaciones humanas recientes, como la exposición a los
rayos X con fines médicos, los materiales radiactivos y las mutaciones
producidas por compuestos químicos, son responsables de lesiones a Nivel de
ADN, bien sea con fines de mejoramiento en lineas celulares de investigación o
simplemente de carácter involuntario por estar expuesto a una fuente de
mutagenos.
Según el mecanismo que ha provocado el cambio en el material genético, se suele hablar de tres tipos de mutaciones: mutaciones cariotípicas o genómicas, mutaciones cromosómicas y mutaciones génicas o moleculares. En el siguiente cuadro se describen los diferentes tipos de mutaciones y los mecanismos causales de cada una de ellas.
| Por sustitución de bases | |||||||||||||||||||||||||
| molecular | |||||||||||||||||||||||||
| Por inserciones o deleciones de bases | |||||||||||||||||||||||||
| Inversiones | |||||||||||||||||||||||||
| Mutación | cromosómica | Deleciones o duplicaciones | |||||||||||||||||||||||
| Translocaciones | |||||||||||||||||||||||||
| Poliploidía | |||||||||||||||||||||||||
| genómica | |||||||||||||||||||||||||
| Aneuploidía | |||||||||||||||||||||||||
Una mutación es el cambio en la
secuencia del DNA, un cambio heredable en el material genético de
una célula.
Aunque la replicación del ADN es
muy precisa, no es perfecta. Muy rara vez se producen errores, y el ADN nuevo
contiene uno o más nucleótidos cambiados. Un error de este tipo, que recibe el
nombre de mutación, puede tener lugar en cualquier zona del ADN.Si esto se produce en la secuencia de nucleótidos que codifica un polipéptido particular, éste puede presentar un aminoácido cambiado en la cadena polipeptídica. Esta modificación puede alterar seriamente las propiedades de la proteína resultante. Por ejemplo, los polipéptidos que distinguen la hemoglobina normal de la hemoglobina de las células falciformes difieren sólo en un aminoácido. Cuando se produce una mutación durante la formación de los gametos, ésta se transmitirá a las siguientes generaciones.
La mayoría de las mutaciones genéticas son perjudiciales para el organismo que las porta. Una modificación aleatoria es más fácil que deteriore y que no mejore la función de un sistema complejo como el de una proteína.
Por esta razón, en cualquier momento, el número de sujetos que portan un gen mutante determinado se debe a dos fuerzas opuestas: la tendencia a aumentar debido a la propagación de individuos mutantes nuevos en una población, y la tendencia a disminuir debido a que los individuos mutantes no sobreviven o se reproducen menos que sus semejantes..
Mutación somática: Es ua mutación que ocurre en tejidos somaticos.
Si esta ocurre en tejido en desarrollo crea una población de celulas mutantes,
con lo que a menudo se expresa fenotipicamente como un sector mutante.
Puede una mutación somática
trasmitirse a la descendencia? Por definición, esto es imposible, sin embargo
hay que tener en cuenta que si se toma un explante de un tejido mutante
donador, y se hace crecer, la planta derivada de el., puede desarrollar tejido
germinal y trasmitir el gen mutante.
Es aquella que ocurre en tejidos
germinales, específicamente en gametos (células sexuales). Si estas células
participan en la fecundación, la mutación se trasmitirá a la siguiente
generación
son aquellas que ocurren a nivel
de un par de nucleotidos en la doble cadena de DNA, puede tener causas
espontáneas o ser inducida.
b) Transversiones: ocurre cuando una pirimidina es sustituida por una purina y viceversa.
c) Mutaciones de cambio de fase: ocurre cuando hay una deleccion de una base, lo que ocasiona que el cuadro de lectura del DNA cambie, esto conlleva a proteinas muy modificadas.
En
cambio, si se omitiera la A (por una deleccion) del codón de iniciación, hay un
cambio de fase de los tripletes, sería el siguiente: UGG, CCU, GUA y ACG, los
cuales se traducen en los aminoácidos triptófano, prolina, valina y treonina,
respectivamente.
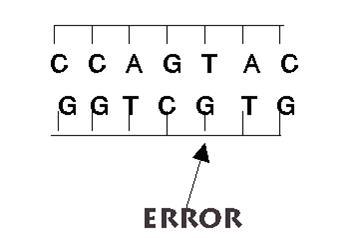
Figura: Mutaciones cambio en la estructura del ADN. A pesar de todos los sistemas destinados a prevenir y corregir los posibles errores, de vez en cuando se produce alguno en la réplica, bien por colocarse una Citosina (C) en lugar de una Timina (T), o una Adenina (A) en lugar de una Guanina (G); o bien porque el mecanismo de replicación se salta algunas bases y aparece una "mella" en la copia. O se unen dos bases de Timina, formando un dímero.


2. Mutaciones
cromosómicas
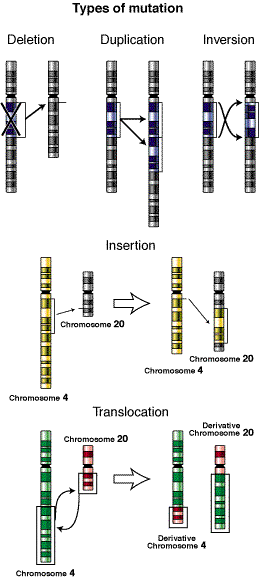
Una parte del cromosoma se puede
separar, invertir y después unirse de nuevo al cromosoma en el mismo lugar. A
esto se le llama inversión. Si el
fragmento separado se une a un cromosoma distinto, o a un fragmento diferente
del cromosoma original, el fenómeno se denomina translocación. Algunas veces se pierde un fragmento de un cromosoma
que forma parte de una pareja de cromosomas homólogos, y este fragmento es
adquirido por el otro.
Entonces, se dice que uno presenta una deleción o deficiencia (dependiendo si
el fragmento que se pierde es intersticial o terminal, respectivamente) y el
otro una duplicación.
Por lo
general, las deficiencias o deleciones son letales en la condición
homocigótica, y con frecuencia las duplicaciones también lo son. Las
inversiones y las translocaciones suelen ser más viables, aunque pueden asociarse
con mutaciones en los genes cerca de los puntos donde los cromosomas se han
roto. Es probable que la mayoría de estos reordenamientos cromosómicos sean la
consecuencia de errores en el proceso de sobrecruzamiento.
Las delecciones de regiones cromosomitas
especificas en humanos producen síndromes únicos, por ejemplo el de “cri du
chat” (grito de gato), cuya causa es una deleccion en el brazo corto (p) del
cromosoma 5, el rasgo característico es el llando semejante al maullido de un
gato de los niños, presentan microcefalia y cara en forma de luna y retraso
mental.


b. cambios en cromosomas enteros y series de
cromosomas
Por ej. En humanos x= 23.
En algunas plantas los monoploides
se pueden obtener artificialmente y después duplicar su numero cromosómico con
colchicina, asi se tendran plantas homocigoticas!
Poliploides
Se deben distinguir entre los Autopoliploides, que se componen de
multiples series de una misma especie y los Alopoliploides que estan compuestos por series procedentes de
diferentes especies.
Generalmente son autopoliploides,
se producen por el cruzamiento entre un 4x (tetraploide) un 2x (diploide). Los
gametos 2x y x se unen y dan lugar a un 3x.
Los triploides son típicamente
esteriles (el problema es por el emparejamientos de cromosomas durante la
meiosis).
Los platanos comerciales son
triploides, con 11 cromosomas en cada serie. La probabilidad de que un gameto
de este 3x contenga exactamente un cromosoma de cada serie es de 1/2048, y hay
una posibilidad en 5 millones que se unan dos gametos de este tipo.
Técnicamente no son esteriles,
solo una fertilidad reducida.
Aparecen naturalmente por una
duplicación espontánea y accidental del genomio 2x a uno 4x, y tambien
artificialmente atravez de la colchicina. Las plantas cosechables autotetraploidesofrecen ventajas,
desde un punto comercial, ya que como suele con los poliploides las series cromosomitas
extras estan asociadas a un incremento de tamaño. Pueden se fértiles o no.
El fenómeno de la poliploidía es
mas comun en plantas que en animales, no obstante hay muchos casos de animales
poliploides. Se encuentran ejemplos en planarias, sanguijuelas y camarones de
mar. (partenogenicos).
Los anfibios y reptiles
poliploides son frecuentes, algunos peces tambien son poliploides, de hecho la
familia de los salmonidos (salmon) aparecio copmo un hecho de poliploidía.
*ANEUPLOIDIA: el número de uno o mas cromosomas pueden cambiar
durante la formación de un nuevo organismo. Dicho organismo se denomina
aneuploide, que significa no-euploide. Puede ocurrir que se añadan cromosomas,
de manera que se tenga un cromosoma extra o que sean eliminados.
Monosomicos (2n-1): cuando se pierde un cromosoma. En tales casos el complemento es perjudicial, por dos razones: los cromosomas que faltan alteran gravemente el equilibrio cromosomico, que ha sido cuidadosamente establecido por la evolución y que es necesario para el equilibrio celular, y la segunda razon, la ausencia de un cromosoma trae como resultado que cualquier alelo recesivo letal situado en el cromosoma sin pareja se exprese directamente.
En humanos la
monosomia para el cromosoma sexual x (44
autosomas y 1 X) produce un fenotipo conocido como síndrome de Turner. Los
afectados presentan unas caracteristicas distinguibles: son hembras esteriles,
de estatura baja, repliegue membranoso en el cuello, inteligencia un poco mas
baja de lo normal, uñas de la mano pequeña, senos poco desarrollados y ausencia
de mestruacion. Los monosomicos para cualquier otro cromososma mueren en el
utero.
En humanos, se conocen varios ejemplos de trisomicos viables. La combinación XXY (1 de 1000 nacimientos de varones) produce el síndrome de Klinefelter: varones altos de aspecto desgarbado, con cierto retraso mental y esteriles, ademas poseen barba poco desarrollada, desarrollo mamario y fisico ligeramente feminizado.
En humanos solo se conocen otros
dos tipos trisomicos en autosomas que sobreviven hasta el nacimiento (en el
cromosoma 13 “síndrome de Patau” y en el cromosoma 18 “síndrome de Edwards”)
ambas trisomias producen graves anormalidades fisicas y mentales. El fenotipo
generalizado de la trisomia en el 13 incluye labio leporino, cabeza pequeña y
deforme, pies arqueados y una esperanza de vida de 130 dias. El fenotipo de los
trisomicos en el 18 incluye posición baja de las orejas, mandibula pequeña,
pelvis estrecha y pies arqueados, mueren en las primeras semanas después del
nacimiento.
En los
siguientes esquemas, tenemos las trisomías más frecuentes tanto en los
autosomas, como en los cromosomas sexuales.
ALTERACIONES EN LOS AUTOSOMAS
SÍNDROME
|
TIPO DE MUTACIÓN
|
Características y síntomas de la mutación
|
Síndrome de Down
|
Trisomía 21
|
Retraso mental, ojos oblicuos, piel rugosa, crecimiento
retardado
|
Síndrome de Edwars
|
Trisomía 18
|
Anomalías en la forma de la cabeza, boca pequeña,
mentón huido, lesiones cardiacas.
|
Síndrome de Patau
|
Trisomía 13 ó 15
|
Labio leporino, lesiones cardiacas, polidactilia.
|
ALTERACIONES EN LOS CROMOSOMAS SEXUALES
Síndrome de Klinefelter
|
44 autosomas + XXY
|
Escaso desarrollo de las gónadas, aspecto eunocoide.
|
Síndrome del duplo Y
|
44 autosomas + XYY
|
Elevada estatura, personalidad infantil, bajo
coeficiente intelectual, tendencia a la agresividad y al comportamiento
antisocial.
|
Síndrome de Turner
|
44 autosomas + X
|
Aspecto hombruno, atrofia de ovarios, enanismo.
|
Síndrome de Triple X
|
44 autosomas + XXX
|
Infantilismo y escaso desarrollo de las mamas y los
genitales externos.
|
5.1.2.2 Agente
mutagénicos
En este grupo encontramos las
radiaciones ionizantes, el calor y recientemente se ha mencionado la exposición
a campos electromagneticos de gran potencia.
Los rayos ultravioleta, gamma y
X, y las emisiones α y β pueden causar mutaciones, por ejemplo los UV ocasionan
daños moleculares y las radiaciones mas fuertes tienden a romper las hebras de
ADN. La radiación cosmica y recientemente el Radon se ha encontrado que son
mutagenicos.
Los mutagenos químicos entran en
tres grupos:
Analogo de bases. Son estructuras similares a las bases
nitrogenadas, pero contienen modificaciones que aumentan la posibilidad de
apareamientos erroneos y tautomerizacion. P.e. 5-Bromuroacilo (5Btu), se
incorpora en lugar de timina, pero tiende a cambiar de forma y aparearse con
guanina..
La 2-aminopurina (2AP) se
comporta como Adenina, pero con frecuencia se tautomeriza y se aparea con la
Citosina.
Agentes desaminantes o alquilantes: estos quimicos modifican los
grupos laterales de bases. Esta modificacion no es en si una mutación pero
induce errores en la replicación. P.e. metil sulfonato, etilmetano sulfonato, dimetilsulfonato,
dimetilsulfato, dietilo sulfato, N-metil-N-nitro-N-nitrosoguanidina, mostaza de
nitrogeno, oxido de etileno.
Desaminacion: tres de las bases del DNA
tienen grupos aminos y estos pueden eliminarse por reaccion con agentes como el
acido nitroso. Los productos de la reaccion y sus propiedades de apareamiento
son:
Adenina
----Hipoxantina, que se aparea con Citosina
Guanina
----xantina, que se aparea con Citosina
Citosina----Uracilo,
que se aparea con Adenina
Alquilacion: diversas posiciones de la
pirimidina son susceptibles a la alquilacion, que producen tanto trasversiones
como transiciones.
Mutagenos que
provocan desplazamiento del marco de lectura:
Se trata de productos quimicos, en especial derivados de acridina, que inducen la inserción o
deleccion de una base, mas que una transición o transversion.
Tanto por daños físico-ambientales
como por errores de síntesis, las biomoléculas pueden sufrir alteraciones
químicas. Puesto que el DNA no puede «recambiarse» como otras biomoléculas ya
que permanece intacto de una división a otra, la estabilidad de la molécula se
consigue mediante dos maquinarias: la fidelidad de la replicación y la reparación
de daños. Los mecanismos de reparación se van a clasificar en 5 grupos
principales:
1. Reparación
directa
2. Reparación
por escisión de nucleótidos (REN)
3. Reparación
por escisión de la base (REB)
4. Reparación
de apareamientos erróneos (mismatch)
5. Sistemas
de recuperación: reparación por recombinación y respuesta SOS
Los agentes que causan daños en el
DNA tienen distintos orígenes. Por una parte, están las alquilaciones
(metilaciones principalmente), las desaminaciones, la oxidación y los rayos UV.
La acumulación de mutaciones en células somáticas es el origen de muchos
cánceres y las células cancerosas reparan mal las mutaciones. Por eso muchos
tratamientos antineoplásicos sobre la base de inducir mutaciones que los
tumores no saben reparar.
CONCLUCIÓN FINAL.
REPARACIÓN DE LOS DAÑOS EN EL ADN
Como hemos venido viendo hasta el momento,
existen muchos agentes físicos y químicos que pueden producir lesiones en el
ADN.
Por tanto, deben existir mecanismos que permitan prevenir y reparar los
daños que se producen en el material hereditario tanto de forma espontánea como
los inducidos.
Como ya hemos visto cuando hablamos de la
replicación del ADN, se dice que la propia ADN polimerasa III posee la subunidad ε que tiene
una función correctora de pruebas que permite detectar cuando EL ADN polimerasa
introduce un nucleótido que no es el correcto y retirarlo.
Este es un
primer mecanismo que evita que se produzcan mutaciones durante la replicación.
Además de este mecanismo existen otros que previenen posibles daños y que
reparan las lesiones producidas:
Superóxido dismutasa: este enzima
convierte los radicales superóxido en peróxido de hidrógeno.
Catalasa: este enzima convierte el
peróxido de hidrógeno en agua.
Gen mutT: este gen codifica para
un enzima que impide la incorporación de la 8-oxo-G al ADN. Este enzima
hidroliza el trifosfato de la 8-oxo-G a la forma monofosfato.
Fotorreactivación: sistema de
reparación directa de los daños producidos por la luz UV. La luz UV produce
dímeros de pirimidinas, fundamentalmente dímeros de Timinas. El enzima
Fotoliasa codificada por el gen phr reconoce en la oscuridad los
dímeros de Timina y se une a ellos, y cuando se expone a la luz (mediante un
fotón) deshace el dímero de Timinas.
Transferasa de grupos alquilo (metilo o
etilo): elimina los gupos alquilo producidos por el EMS o por NG. El enzima
metiltransferasa transfiere el grupo metilo de la O-6-metilguanina a una
cisteína (cys) de la enzima.
Reparación de los daños de la luz UV
(Endonucleasa uvrABC): La Endonucleasa uvrABC es una
escilnucleasa codificada por los genes uvrA, uvrB y uvrC que corta el
ADN. La Helicasa II de ADN separa las dos hélices y retira 12
nucleótidos. La ADN polimerasa I rellena el hueco producido por la
Helicasa II y la Ligasa sella los extremos.
Reparación AP: reparación de las sedes
apurínicas o apirimidínicas. La llevan a cabo las Endonucleasas AP de la
clase I que cortan por el extremo 3' y las de la clase II que cortan por el
extremo 5'. Una exonucleasa elimina una pequeña región que contiene entre
dos y 4 nucleótidos, la ADN polimerasa I rellena el hueco y la
Ligasa sella los extremos.
Reparación mediante glucosidasas: estas
enzimas detectan las bases dañadas y las retiran rompiendo el enlace
N-glucosídico con el azúcar. Como consecuencia se origina una sede AP que se
repara de la forma indicada anteriormente (reparación AP). La Glucosidasa de
Uracilo elimina el Uracilo (U) del ADN.
La Glucoxidasa de
Hipoxantina, elimina la Hipoxantina (H) del ADN. Además de estas dos
glucosidasas existen otras diferentes.
Sistema GO: dos glucosidasas producto
de los genes mutM y mutY actúan conjuntamente para eliminar las
lesiones que produce la 8-oxo-G (GO).
Reparación de apareamientos
incorrectos: la reparación de apareamientos incorrectos posterior a la
replicación requiere la existencia de un sistema que sea capaz de realizar las
siguientes operaciones:
- Reconocer las bases mal apareadas.
- Determinar cual de las dos bases es la incorrecta.
- Eliminar la base incorrecta y sintetizar.
Esta reparación la realizan los productos de
los genes mutH, mutL, mutS y mutU. Además, para distinguir la hélice de nueva
síntesis de la hélice molde y así saber eliminar la base incorrecta, el sistema
consiste utiliza el hecho de que la hélice de nueva síntesis tarda un cierto
tiempo en metilarse la Adenina (A) de la secuencia GATC, mientras que la A de la
secuencia GATC de la hélice molde ya está metilada. El enzima que reconoce la
secuencia GATC metilando la A que contiene es la Metilasa de
Adenina.
 |
 |
| Reparación directa: Fotorreactivación | Reparación por escisión: daños UV |
 |
 |
| Reparación posterior a la replicación | Reparación sedes AP |
Reparación por recombinación:
ademas se dice que cuando el
ADN polimerasa III encuentra un dímero de Timina (T) producido por luz UV no
sabe que nucleótido poner saltando la región y dejando un hueco.
Como
consecuencia esa región queda como ADN de hélice sencilla. Debido a que la luz
UV produce muchos dímeros de Timina, se produce un bloqueo en la replicación y
para evitar que la célula muera y pueda replicarse se dispara el sistema de
emergencia SOS. La puesta en marcha del sistema SOS comienza porque el ADN de
hélice sencilla activa a la proteína RecA que a su vez interacciona con
la proteína LexA.
La proteína LexA es un represor de los genes
uvrA, uvrB, uvrD, sulA y sulB. Todos estos genes tienen en el promotor
una secuencia denominada caja SOS. La proteína LexA normalmente impide la
transcripción o expresión de los genes citados anteriormente, pero cuando
interacciona con la proteína RecA deja de impedir la expresión de estos
genes, pudiéndose sintetizar las proteínas correspondientes y reparar los daños
producidos por la luz UV.
Restauración total o parcial del fenotipo
normal (apariencia externa, actividad proteica) a partir de un individuo
mutante.
Reversión genotípica: se produce por
causas genéticas.
Reversión fenotípica: se produce por
causas no genéticas (ambientales).
La reversión es una 2ª mutación que restaura
total o parcialmente el fenotipo normal a partir de un individuo mutante. Al
individuo que recupera el fenotipo normal por medio de esta 2ª mutación se le
denomina Revertiente.
REVERSIÓN GENOTÍPICA
La reversión genotípica se puede conseguir de
dos maneras:
- Retromutación (en sentido estricto): consiste en recuperar la secuencia exacta de nucleótidos en el ADN.
- Mutación supresora: es una mutación secundaria que restaura total o parcialmente la función pérdida.
RETROMUTACIÓN
Consiste en recuperar la secuencia exacta de
nucleótidos en el ADN, un ejemplo de esta situación sería: AAA
(lys)→GAA(glu)→AAA(lys).
A veces también se habla de retromutación
en sentido funcional, de manera, que se recupera la actividad enzimática
normal pero no se recupera la secuencia original de nucleótidos en el ADN, por
ejemplo: UCC(ser)→UGC(cys)→AGA(ser). Sin embargo, el concepto de retromutación
funcional no se diferencia del concepto de mutación supresora y puede inducir a
error.
http://www.ucm.es/info/genetica/grupod/Mutacion/mutacion.htm#Evitan errores
http://es.wikipedia.org/wiki/Reparaci%C3%B3n_del_ADN
http://es.wikipedia.org/wiki/Mutaci%C3%B3n
No hay comentarios:
Publicar un comentario