SEP GNEST DGEST
INSTITUTO TECNOLOGICO DE CD ALTAMIRANO

UNIDAD 6. TRANSCRIPCIÓN
GENÉTICA.
ALUMNO: CANDIDO TORRES SANTIBAÑEZ
CON # DE CONTROL; 09930058.
PROFESOR; FRANCISCO JAVIER PUCHE ACOSTA.
CD ALTAMIRANO,GRO.MEXICCO.
TEMAS DE LA UNIDAD # 6
6.1 Organismos procarióticos:
6.1.1 Etapas de síntesis del ARN.
6.2 Organismos eucarióticos:
6.2.1 Etapas de síntesis del ARN.
6.2.2 Modificaciones postranscripcionales del RNA
mensajero INTRODUCCION.
Qué es la transcripción?
La transcripción del ADN es
el primer proceso de la expresión génica,
mediante el cuál se transfiere la información contenida en la secuencia del ADN
hacia la secuencia de proteína utilizando
diversos ARN como intermediarios. Durante la
transcripción genética, las secuencias de ADN son
copiadas a ARN mediante una enzima llamada ARN polimerasa que sintetiza un ARN mensajero que mantiene la
información de la secuencia del ADN. De esta manera, la transcripción del ADN
también podría llamarse síntesis del ARN mensajero.
a transcripción consiste en la copia de 1 cadena de DNA para dar
una cadena de RNA, gracias a la complementariedad de bases. La cadena que se
copia se conoce como cadena molde o cadena transcrita. Esta cadena será
obviamente complementaria al RNA.
Se conoce como unidad de transcripción a aquel DNA
que da lugar mediante el proceso de transcripción a una molécula de RNA. No
siempre se corresponderá una unidad de transcripción con un gen, ya que en los
organismos procariotas podrán existir operones, con lo que se coordinarán
varios genes, que se transcribirán de manera simultánea.
Para la transcripción resulta básica la presencia
de la RNA polimerasa, ya que este enzima es el encargado de sintetizar el RNA,
gracias a la complementariedad con la cadena molde. La RNA polimerasa da inicio
a la transcripción cuando se une al promotor. Se conoce como punto +1 al punto
donde se inicia la transcripción. El enzima se deslizará por el molde hasta
alcanzar la secuencia acabadora, situada en la parte final de la unidad de
transcripción. Todos los nucleótidos situados antes de +1 son los situados
upstream o hacia 5’. Estos nucleótidos reciben numeración negativa. Los
situados después de +1 están situados downstream o hacia 3’.
A lo largo del DNA, las unidades de transcripción
pueden situarse en cualquiera de las dos cadenas, lo que implicaría
dificultades para ilustrar este proceso. Se ha determinado arbitrariamente, que
la transcripción se inicie siempre de izquierda a derecha, desde 3’ a 5’. En
paralelo a la cadena de RNA se pone una sola cadena de DNA, pero no la
complementaria, sino la que es idéntica al RNA, con las diferencias típicas
entre DNA y RNA, como la sustitución de T por U.
El RNA que se forma como resultado de la
transcripción, podrá ser el tránscrito primario. Se ha de tener en cuenta que
existe tres tipos de RNA, dos de los cuales son productos finales, como el rRNA
y el tRNA, mientras que el mRNA deberá llegar a los ribosomas, donde podrá dar
lugar a las proteínas.
Dentro del proceso de la transcripción puede haber
otras proteínas implicadas, que serán las proteínas reguladoras. Se ha de tener
en cuenta que la mayoría de los genes está sometidos a regulación, de manera
que existirán diferentes ritmos de síntesis de RNA, con más o menos frecuencia.
Esta regulación afecta a la expresión del gen, actuando a nivel de la
transcripción normalmente.
Muchas de las cosas anteriormente mencionadas se
pueden aplicar tanto a procariotas como a eucariotas, por lo que nos
centraremos a partir de este punto en la transcripción en procariotas.
El caso que estudiaremos más el de la bacteria
E.coli, que posee unos 5000 genes. Al tratarse de una bacteria, posee un único
cromosoma, circular. La regulación se da principalmente a nivel de la
transcripción. Siendo una bacteria, posee un sistema de rgulación típico en las
bacterias, que es la coordinación de diferentes genes mediante una estructura
característica de las bacterias, que se conoce como operones. Los operones
permiten la coordinación de diferentes genes bajo un mismo promotor, dando como
resultado un único RNA. La inmensa mayoría de los organismos procariotas carece
de intrones repartidos a lo largo de su genoma.
La RNA polimerasa es el único enzima de síntesis
de RNA, capaz de sintetizar los 3 tipos diferentes de RNA. Existirán 2 tipos de
promotores, los fuertes, con un alto índice de transcripción, y los débiles,
con una transcripción más reducida. Estos promotores dependen de su afinidad
por la RNA polimerasa.
OBJETIVO GENERAL DEL CURSO
Entender las bases moleculares del control de la expresión génica y de la manipulación del
ADN mediante el empleo de las diferentes técnicas moleculares que le permitan
desarrollar proyectos de investigación básica y aplicada.
OBJETIVO DE LA UNIDAD # 6
· Conocer los eventos de síntesis del ARN como
molécula precursora de la síntesis proteica y su importancia en el
funcionamiento de los seres vivos.
METODOLOGIA
La
metodología que se utiliza son los pasos a seguir para la manifestación y desenlace de la unidad # 6 y con sus correspondientes subtemas: realizando enlaces de acuerdo a lo manifestado en clases de la unidad por el maestro y hacer conclusiones a lo entendido al final
de la unidad de todos los subtemas, publicando lo más interesante yde cada tema visto en clase. y trabajos o ensayos obtenidos correspondiente a cada clase.
UNIDAD 6. TRANSCRIPCIÓN GENÉTICA

6.1 ORGANISMOS PROCARIOTICOS:
La formación
de una cadena de ARNm por una secuencia particular de ADN se denomina
transcripción. Antes de que termine la transcripción, el ARNm comienza a
desprenderse del ADN.
La expresión génica se concretiza por la transformación de la información
genética desde moléculas de ADN a moléculas de ARN y desde estas hasta los poli
péptidos correspondientes. Las moléculas de ARN son sintetizadas usando
como molde a segmentos específicos de ADN, en la reacción de polimerización que
es catalizada por la enzima conocida como ARN polimerasa.

La trascripción utiliza una de las cadenas
del DNA como molde
La
trascripción depende de la complementariedad entre las bases. Se produce una
separación local de las dos cadenas del DNA, y una de las dos cadenas actúa
como molde (guia) para la síntesis
de RNA. A lo largo del DNA los genes utilizan una u otra cadena para copiarse
aRNA (pero un gen especifico siempre
usara una cadena y solo una como molde).
A
continuación, nucleotidos libres se emparejan con el DNA molde “atraidos” por
las bases complementarias. El nucleotido A empareja con T en el DNA, G con C, C
con G y U con A. Este proceso esta catalizado por la polimerasa de RNA, que se une al DNA y se mueve a lo largo de la
molécula añadiendo ribonucleótidos al RNA naciente.
El
crecimiento del RNA se produce siempre en la direccion 5´ 3´
Polimerasas de RNA
En las
procariotas una sola enzima polimerasa trascribe todos los RNAs, pero en
eucariota setas especializadas asi:
- polimerasa
de RNA I (Pol I) trascribe los genes que determinan RNAr.
- la
polimerasa de RNA II (Pol II) trascribe los genes que determina proteinas,
o lo que es lo mismo RNAm.
la polimerasa de RNA III (Pol
III) trascribe otros RNA funcion
Promotores bacterianos
La secuencia
reconocida por la RNA-polimerasa en el DNA para comenzar la transcripción se
denomina promotor. Sus características estructurales fueron resueltas en 1975
por David Pribnow y Heinz Schaller de manera independiente (téngase en cuenta
que la secuenciación se estandarizó en 1977). Presentan una secuencia común
rica en A y T en la posición -10 (caja TATA o Pribnow); la presencia de pares
AT en esta caja permite la separación fácil de las dos cadenas de DNA. Esta es
la secuencia que reconoce específicamente la subunidad σ. A -35 se encuentra
otra secuencia conservada cuyo consenso es TTGACA..
Los nucleótidos de la región
-10 y -35 son los principales puntos de contacto entre la proteína y el DNA. La
distancia óptima entre las cajas es de 17 nt, lo que equivale a dos vueltas de
hélice, por lo que la RNA-polimerasa se une al promotor sólo por un lado del
DNA, contactando siempre la misma cadena.
6.1.1 ETAPAS DE SINTESIS DEL ARN.

Iniciación
Se puede
subdividir en varias etapas: en cuanto la holoenzima RNA-polimerasa se
encuentra con la secuencia promotora, se forma el complejo promotor cerrado y
la afinidad de esta unión es muy alta. La zona que la enzima protege en el DNA
abarca desde +20 hasta -55. A continuación la propia RNA polimerasa desenrolla
12 pb del DNA desde -10 hasta -1 (complejo promotor abierto).
Mediante una
isomerización dependiente de Mg2+, la separación de las cadenas se extiende
desde -12 hasta +2 y el DNA se curva fuertemente en forma de U en la
RNA-polimerasa. Se necesitan las topoisomerasas I y II para controlar el
superenrollamiento en las zonas de transcripción, al igual que ocurre durante
la replicación. El primer nucleótido lo coloca el centro activo que tiene más
afinidad por A o G (por eso uno de ellos es el primer nucleótido de la mayoría
de los transcritos).
Si la transcripción no se ha abortado al 10º nt, entonces
se disocia la subunidad σ del complejo de transcripción, se pierde el contacto
con la zona de -55 a -30, y se considera terminada la etapa de iniciación. La
disociación de σ es absolutamente necesaria para que se pueda empezar a
desplazar la polimerasa central.
Elongación
Basta la
polimerasa central para realizar toda la etapa de elongación. La burbuja de
transcripción abarca 18 nt de los que 8-9 nt forman un heteroduplex con los
últimos nt transcritos del RNA (extremo 3’).
El avance de
la polimerasa no es continuo, sino a saltos, por lo que se denomina avance en
forma de gusano. Este avance en gusano se ve, además, retrasado debido a que
durante la elongación se producen numerosas paradas porque hay secuencias que
se transcriben peor que otras. En otras ocasiones puede deberse a que se ha
producido un error durante la transcripción. En estas paradas, la
RNA-polimerasa retrocede unos nucleótidos, de manera que el extremo 3’ del RNA
naciente queda protuberante y sin aparear, lo cual impide que se pueda seguir
transcribiendo.
Cuando la
RNA-polimerasa se acerca a la región terminadora, pasa sobre una secuencia
denominada nut que sirve para
incorporar a la polimerasa central la subunidad NusA. Inicialmente se creía que
NusA serviría para disminuir la afinidad de la polimerasa por el DNA, pero
parece que lo que provoca es un aumento inespecífico de paradas durante la
polimerización, lo que facilita la terminación de la transcripción.

Terminación intrínseca
También se
denomina —independiente de Ro—. Necesita dos regiones ricas en GC simétricas y
complementarias con lo que se puede formar una horquilla, seguidas de una
región rica en A. Al llegar a las secuencias ricas en GC, la RNA-polimerasa con
NusA reduce su velocidad, dando tiempo para que el transcrito pueda formar la
horquilla y deshaciendo la interacción con HBS. Cuando la enzima pasa sobre la
secuencia rica en A, el híbrido DNA-RNA es muy débil y se disocia. Las
secuencias anteriores y posteriores a la horquilla y oligo(A) también influyen,
pero no se sabe cómo.

Es menos
abundante en los organismos. Necesita el factor de terminación Ro
(NusD) de 46 kDa que actúa como homohexámero (275 kDa). hace separarse
el RNA naciente, la polimerasa y el DNA.

6.2 ORGANISMOS EUCARIOTICOS:
La
estructura de los ARNm y el proceso de transcripción en las células eucariotas,
es similar a lo expresado anteriormente para las células procariotas. Sin
embargo, debemos considerar las siguientes diferencias:
a) Las células eucariotas poseen tres clases de ARN polimerasas (I, II y
III), las cuales se utilizan para sintetizar los distintos tipos de ARN
existentes.
b) Los extremos 3’ y 5´ de los ARNm están modificados. En el caso del
extremo 5’ encontraremos una estructura denominada CAP y, en el
correspondiente extremo 3’, se encuentra adherido una larga secuencia de
nucleótidos cuya base nitrogenada es la Adenina (Cola Poly A).
c) Las moléculas de ARNm, luego de ser sintetizas, son modificadas. Los
“transcriptos primarios” eucariotas sufren un proceso por el cual determinadas
secuencias (Intrones) son eliminadas.
d) Los ARNm eucariotas son Monocistrónicos.
Buena parte
de las peculiaridades de la transcripción eucariótica se debe a que estas
células producen más tipos de RNA que los procariotas, lo que lleva a una clara
distinción entre los que es un transcrito primario (o RNA precursor) y
un transcrito maduro que será el RNA funcional. En los eucariotas, todos
los transcritos primarios sufren algún tipo de maduración o procesamiento.
Localización intracelular.
La
localización intracelular también va a ser específica: en una célula eucariota,
por ejemplo un fibroblasto, encontramos aproximadamente 6 pg de DNA que
corresponde a su material genético. En cambio, encontramos 25 pg de RNA, de los
que 3-4 pg están localizados en el núcleo y el resto en el citoplasma.
Todos
los transcritos primarios y algunos de los maduros (por ejemplo los snRNA,
véase más adelante) se encuentran en el núcleo, mientras que en el citoplasma
sólo se encontrarán formas maduras de la mayoría de los transcritos. De los que
se encuentran en el núcleo, los pre-rRNA se encuentran en el nucléolo y los
demás en el nucleoplasma.

Las tres
RNA-polimerasas eucarióticas codificadas en el núcleo se numeran I, II y III en
función del orden en el que eluyen cuando se cromatografía un extracto nuclear
en columna de intercambio iónico al ir aumentando la fuerza iónica del tampón
de elución.
Las tres se transcriben desde distintos genes, se localizan en
diferentes compartimentos, son susceptibles a distintos inhibidores, tienen un
tamaño en torno a 600 kDa y están compuestas por dos subunidades grandes no
idénticas que se denominan, como en procariotas, núcleo polimerasa o polimerasa
central.

LOS EXALTADORES O ENHACEARS
Los promotores Eucariotas no trabajan solos para asegurar una
transcripción eficaz. En algunos genes o tipos celulares, la actividad de un
promotor está aumentada por la presencia de otra secuencia conocida como
Exaltador o Enhacer.
Las características de los Exaltadores son las siguientes:
• Están formados por cientos de bases
• Generalmente incluyen secuencias repetidas
• Actúan a distancia, miles de bases del promotor.
• Son activos en cualquier orientación respecto al promotor,
incluso suele encontrárselos dentro del mismo gen.
Los exaltadores, entonces, actúan como reguladores de la expresión
génica.
No está claro aún el mecanismo de acción de los exaltadores, más aún
cuando estos se encuentran localizados a cualquier distancia (1000 a 2000
Pb) y dirección del mencionado promotor.

El ARN mensajero en Eucariotas:
Si bien la síntesis y estructura de los ARN mensajeros en Eucariotas es
similar a las Procariotas, deben mencionarse las siguientes diferencias:
a) Ambos extremos están modificados, el 5’ presenta una estructura
denominada CAP (caperuza) y el 3’ contiene una secuencia de Ácido Poliadenílico
denominada Cola Poly A.
b) La molécula que sirve de molde para la síntesis de proteína
(ARNm maduro) es más corta que el ARN mensajero recién sintetizado (Pre –
ARN mensajero). Esto se debe a que los últimos sufren un proceso de eliminación
de secuencias no codificantes. Dicho proceso se denomina Splicing.

6.2.1 ETAPAS DE SINTESIS DEL ARN.
Al igual que
en procariotas, las etapas básicas son la iniciación, la elongación
y la terminación. Los procesos enzimológicos también son los mismos, por
lo que vamos a ver cuáles son las diferencias esenciales que caracterizan al
proceso eucariótico.
Hasta no
hace mucho, los promotores se analizaban por deleciones seriadas desde el
extremo 5' del gen, por lo que se consideraba que el promotor empezaba a partir
del nucleótido más distal cuya deleción provocaba la pérdida de la capacidad
transcriptora. Siguiendo este criterio, quedaban excluidos del promotor muchos
elementos reguladores que no influían decisivamente sobre la transcripción del
gen.
Para la
transcripción son necesarios una serie de factores de transcripción que
se nombran empezando por la abreviatura TF, le sigue el número de la polimerasa
que reconoce, y termina por una letra que sirve para distinguirlos unos de
otros. Por razones históricas, existen muchas excepciones a esta regla.
Durante la
iniciación es cuando se produce el reconocimiento del promotor y asociación de
la polimerasa. Es la parte más compleja de la transcripción y sobre la que se
ejercerán la mayoría de las actividades reguladoras. Debido al gran tamaño del
genoma, los factores de transcripción y la polimerasa tienen muy difícil
reconocer sus sitios específicos. Para conseguir especificidad se requieren
mecanismos más intrincados como el superenrollamiento —poner en forma de
heterocromatina las regiones de DNA que no se tienen que expresar— con el
objeto de disminuir la cantidad de DNA a rastrear. Por eso, como se verá, la
presencia de los nucleosomas entorpece la transcripción de los genes.

La función
del promotor es, esencialmente, facilitar la unión de la RNA-polimerasa al DNA
y asegurar que el inicio de la transcripción se realice muy eficazmente y en un
nucleótido determinado. El complejo de iniciación produce en la región de
inicio de transcripción el desenrollamiento y separación de las dos cadenas del
DNA (complejo abierto) que es donde va a tener lugar la síntesis de la
cadena de RNA. Esta estructura se desplazará por el DNA a medida que avanza la
elongación (burbuja de transcripción).
Es necesario que se en torno a la
RNA-polimerasa forme el complejo holoenzimático polimerasa para que se
pueda empezar a transcribir. El modelo más aceptado propone que en la región
promotora se van asociando los factores de transcripción y subunidades de la
holoenzima de manera ordenada y constante, hasta que se une la polimerasa
central.
También se requiere la presencia de una DNA-helicasa y la
establización de la región desapareada con proteínas específicas. El proceso
enzimático por el que se añaden los ribonucleótidos uno a uno a partir del
primer nucleótido trifosfato es igual que el procariota.
cada
RNA-polimerasa reconoce un promotor específico y forma un complejo
holoenzimático polimerasa diferente y característico.

Decondensación del DNA para la
transcripción
Para poder
iniciar una transcripción, la maquinaria transcriptora ha de poder acceder a la
hebra de DNA, lo que implica la descondensación previa de la región implicada
del cromosoma.
Durante la elongación de la
transcripción, la RNA-polimerasa es capaz de desplazar los nucleosomas en la
zona codificante: cuando la RNA-polimerasa se encuentra con uno, lo empuja sin
esfuerzo los primeros 30 nt y luego se ralentiza hasta el nt 80 —con paradas
cada 10 nt—, momento en el que el nucleosoma es desplazado por detrás de la
enzima gracias a la energía potencial topológica que se acumula en las
superhélices por la tensión del DNA.
La holoenzima ha
permanecido inmóvil mientras se polimerizan los primeros nucleótidos, hasta que
el TFIIK fosforila el CTD, activado por el TFIIE. Tras la fosforilación, la
polimerasa se libera de todos los factores de iniciación excepto la helicasa
del TFIIF y comienza su avance por el DNA. En torno al elemento Inr pueden
quedarse los TFII A, B y D así como otros activadores de la transcripción,
listos para reiniciar otro ciclo de transcripción —esta vez más abreviado
puesto que las primeras estapas no han de repetirse.
Una
actividad helicasa sigue abriendo el DNA por delante, lo que genera importantes
tensiones en la molécula que se compensan por la acción de las topoisomerasas.
Por detrás de la polimerasa el RNA recién sintetizado se va desapareando del
DNA molde y quedando en forma de cadena sencilla de RNA. Las dos cadenas de DNA
se vuelven a aparear y gracias a la asistencia de otra topoisomerasa, se
rebobina de nuevo el DNA.


En esta etapa intervienen al menos 16 factores de elongación cuyo mecanismo exacto se desconoce. Su función siempre parece relacionada con 3 fenómenos principales:
- la eliminación de las pausas o paradas de la polimerasa. Los mejor identificados son TFIIS (antes llamado SII) que sirve para evitar que la RNA-polimerasa se detenga prematuramente, y los que alteran la Km y la Vmax de la enzima para incrementar su capacidad catalítica: ELL, CSB y SIII (elonguina).
- el mantenimiento del estado fosforilado del CTD (PTEFb y otros), ya que su desfosforilación inhibe la elongación.
- factores que posteriormente intervendrán en el ayuste, así como proteínas que intervienen en la remodelación de los nucleosomas durante la elongación (SWI/SNF, FACT, HMG14, elongator...), ya que sigue siendo necesario despejar el camino.
Terminación: Las secuencias terminadoras propiamente
dichas están aún poco caracterizadas, pero se ha visto que provocan dos
efectos:
- detención o pausa en el avance de la polimerasa al pasar por ellas
- desestabilización del híbrido RNA—DNA.
El extremo 3’ de los RNA eucarióticos transcritos no coincide con el extremo 3’ de los RNA maduros ya que es habitual que los extremos de los transcritos primarios se corten durante la maduración (excepto los de la RNA-polimerasa III). También se comprende la dificultad del estudio de las secuencias y mecanismos de terminación, ya que los transcritos primarios (hnRNA y otros pre-RNA) son de corta vida y difícil purificación.
Terminación de la RNA-polimerasa I
La señal de terminación de los rRNA es una secuencia de 18 pb duplicada que se encuentra triplicada, de la que hay dos copias cerca del extremo 3’ del gen 28S (T1 y T2) y la tercera a 200 pb del inicio de transcripción del gen siguiente (T3).Esta enzima tiene un mecanismo equivalente al de la terminación independiente de ρ en procariotas. En una secuencia indeterminada se une una proteína terminadora (Reb1p en levaduras o TTF-I en mamíferos) que detiene el avance de la RNA-polimerasa. Una vez detenida, una región rica en T en la cadena codificante facilita la separación del RNA. No se ha observado que se formen horquillas.

Terminación de la RNA-polimerasa II
Esta enzima tiene un mecanismo
equivalente al de la terminación dependiente de ρ de procariotas. Se ha
observado que la enzima se detiene en la región de terminación debido bien a
secuencias específicas en el DNA, o bien a proteínas que favorecen la pausa que
aún no se han identificado. Una proteína similar a ρ se une al RNA e induce la
separación del complejo holoenzimático. De hecho se ha comprobado que la propia
ρ de E. coli es capaz de inducir esta terminación. Está directamente
relacionada con la poliadenilación del mRNA.

Terminación de la RNA-polimerasa III
Esta enzima tiene un mecanismo equivalente al de la
terminación intrínseca en procariotas. La detención de la RNA-polimerasa se
produce en alguna de las T que hay al final del gen. Se producen diferencias en
la terminación dependiendo del tipo de promotor que se ha utilizado en la
iniciación:
- Si es un promotor de tipo «a», el TFIIIA atrae los factores de terminación específicos que van a reconocer las T si van precedidas de un palíndromo de GC.
- Si es un promotor de tipo «b» es TFIIIC el que atrae los factores de terminación.
- Si es un promotor de tipo «c», SNAPc es la que va a reclutar los factores de terminación que van a reconocer las T dentro de una zona rica en A puesto que se ha visto que la presencia de G inhibe la terminación en este caso.
A diferencia de la terminación de las otras dos polimerasas,
estos transcritos no van a sufrir ninguna modificación en su extremo 3’, por lo
que todos acaban en varias UUU.

.2.2 MODIFICASIONES POSTRANSCRIPCIONALES DEL RNA MENSAJERO.

Los productos
inmediatos de la transcripción se denominan transcritos primarios y no son
necesariamente funcionales; muchos de ellos son específicamente alterados en
varias formas como son:
1.
Remover segmentos exo o endonucleotidicos.
2.
Adición de secuencias nucleotidicas en cualquiera de los dos
extremos (5´ ó 3´).
3.
Modificación de nucleótidos específicos.
Estas
modificaciones no son iguales para todos los tipos de ARN, cada uno tiene
sus propias particularidades:
En los procariontes, la mayoría de los
transcritos primarios son funcionales inmediatamente después, e incluso durante
su síntesis. Es decir estos mARN participan en la traducción, sin
modificación alguna. De hecho, en estos organismos, los ribosomas usualmente
comienzan la traducción en la cadena naciente del mensajero.
En los eucariontes por
el contario, el mARN es producido en el núcleo y su
traducción ocurre en el citoplasma. En el nucleoplasma, durante el camino
al citoplasma, los mARN sufren alteraciones en su estructura. A estos
cambios se les denomina modificaciones postranscripcionales.
PROCESO:
Los mARNs eucarioticos estan encapuchados

Los mARNs eucariontes maduros, poseen una
capucha que es agregada enzimáticamente y
que consiste de un residuo de 7-metilguanosina unido en el extremo inicial (5´) por
un enlace trifosfato (5´-5´):

LA ESTRUCTURA DE LA CAPUCHA DE LOS MARNS EUCARIONTES SE PUEDE ENCONTRAR EN TRES
FORMAS:
0: no tiene modificaciones (forma predominante
en organismos eucariontes unicelulares).
1: el nucleótido líder está 02´metilado (forma
predominante en organismos multicelulares).
2: los dos primeros nucleótidos están 02´metilados.
La capucha es unida
por una guanililtransferasa específica, después de » 20 nucleósidos,
define el sitio de inicio de la traducción. Si elnucleósido líder es adenosina (generalmente es
una purina), puede estar también metilada en
posición N6.
Los mARNs tienen colas de poliA
Los mARNs eucariontes
a diferencia de los procariontes, son siempre monocistrónicos, esto quiere
decir que contienen información únicamente para un gen, a diferencia de
los operones procariontes.
La señal de terminación de la
transcripción el los eucariontes, no se ha determinado con precisión, esto se
debe a que el proceso es impreciso i.e. los transcritos
primarios de los genes estructurales tienen secuencias 3´
heterogéneas. A pesar de lo anterior, los ARNs maduros
presentan secuencias 3´ definidas, casi todos ellos
terminan en colas de poli A de 20 a 50 nucleótidos, que no son necesarios para
la transcripción in vitro. Las colas de poliA, se unen a
la proteína de unión al poli(A) (PABP), lo que genera
una ribonucleoproteína. PABP protege al mARN; la presencia de esta
proteína reduce la velocidad de degradación del mARN.
La mutación de la
cola poliA de un transcrito presenta una vida media (t1/2)
menor a 30 min en el citoplasma, por el contrario, el
mismo transcrito con su cola de poliA tiene una t1/2 que
varía de horas a días. Esta cola de poliA, es agregada
al transcrito primario en dos reacciones.
1.- el transcrito es cortado en una
posición entre 15 a 25 nucleótidos pasando una secuencia conservada AAUAAA, que
al mutarse, inhibe el corte y la poliadenilación.
2.- la cola de poliA se genera a
partir de ATP, gracias a la catálisis de la poli(A)polimerasa

Los genes eucariontes consisten de secuencias alternadas de expresion y de no expresion.
La mayor diferencia
entre los genes estructurales eucariontes y procariontes es que la secuencias
de la mayoría de los genes eucariontes, combinan secuencias de expresión con
secuencias de no expresión. Los transcritos primarios son muy heterogéneos en
longitud (desde » 2000 hasta más de 20,000 nucleótidos) y son mucho
más largos de lo que se esperaría para el tamaño de las proteínas que
producen.
En 1977 Phillip Sharp y Richard Roberts,
describieron independientemente que los pre-mARNs son procesados por
la escisión de secuencias internas. Los pre-mARNs contienen
típicamente ocho secuencias no codificantes o intrones que
agregan una longitud que varia entre 4 y 10 veces la longitud de las
secuencias codificantes o exones. Para dar una idea de
el descubrimiento de estos investigadores, se presenta la siguiente Figura
en donde se representa a la proteína ovoalbumina de pollo, que por cierto
es la proteína más abundante en la clara de huevo.

Figura: Representación de la alineación de los
segmentos complementarios de ADN (línea azul) y mARN (línea
discontinua roja) del gen de la ovoalbumina de pollo.
Los números arábigos representan la posición
de los exones (1-7). Los números romanos representan las
secuencias complementarios en elmARN i.e. los intrones.
La longitud de los intrones en los
vertebrados varía entre » 65 a » 200,000 nucleótidos con
periodicidad no obvia. La formación delmARN eucarionte comienza con la
transcripción del gen estructural completo (incluyendo los intrones)
formando el pre-mARN, después viene la agregación de la capucha y la cola de poliA, posteriormente, se
cortan los intrones y se pegan los exones, dando origen
al mARN maduro.
Este proceso de edición (splicing en
inglés), se hace con mucha precisión, pues si se deja o corta una base de más,
la proteína producida, puede no ser funcional; los exones nunca se
mueven de lugar, tienen el mismo orden en el mARN maduro que en el
gen.
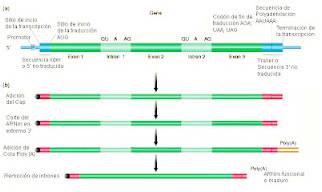
Ayuste de intrones
Durante o
poco tiempo después de la síntesis de los pre-MARNs de vertebrados, 0.1% de
los residuos de A, están metilados en posición
N6. Estos m6As, tienden a ocurrir en secuencia RRm6ACX, en donde X es
raramente una G. La función de este proceso es desconocida, pero una gran
cantidad de estos residuos forman parte del mARN maduro.
EVIDENCIAS
Al terminar cada unidad el profesor aplica y aplicara en esta unidad un examen para poder evaluar la unidad, y en cuestion de trabajos, el profesor nos indica realizar practicas y ensayos correspondientes a la unidad osea de los temas mas importantes y sobre salientes que en caso comun todos los temas y subtemas son de gran interes de estudio.. y ricos en aprendizaje.
CONCLUSIÓN
A paso de estudio de esta unidad # 6 " transcripción genética" se observan y se manifiesta
Diferencias entre la transcripción procariótica y eucariótica que acontinuacion se ve.
Diferencias entre la transcripción procariótica y eucariótica que acontinuacion se ve.
- La cromatina a transcribir está mucho más condensada en los eucariotas: en la metafase (máxima condensación) la transcripción está detenida.
- La mayor parte del DNA eucariota (como mínimo la mitad) no se transcribe nunca, mientras que en los procariotas se transcribe práticamente todo.
- Todos los transcritos eucariotas han de madurarse en el núcleo, aunque algunas etapas pueden ocurrir en el citosol.
- En los eucariotas, la transcripción y la traducción no están acopladas, y se producen en compartimentos diferentes. Esto implica una regulación distinta para cada proceso.
- Las secuencias 5’ y 3’ no traducidas de los mRNA son mucho más largas que las que se encuentran en los procariotas.
- Los genes eucariotas son monocistrónicos.
- En los eucariotas hay 3 polimerasas nucleares distintas, y también una característica de mitocondrias y una quinta para los cloroplastos.
- Las polimerasas eucariotas necesitan muchos factores de transcripción para funcionar. La mayoría son activadores.
- Los eucariotas producen más tipos de transcritos distintos
- La transcripción eucariótica es muy específica y regulada, lo que les confiere una expresión muy controlada, compleja y precisa.
- Se puede conseguir silenciar completamente un gen en un eucariota.
BIBLIOGRAFIA:
http://es.wikipedia.org/wiki/Transcripci%C3%B3n_gen%C3%A9tica
Hola como estás? gran parte de este texto tiene copia de mi Blog. Deberías agregarlo como fuente www.genmolecular.com "Desde Mendel hasta las moléculas"
ResponderEliminar