INSTITUTO TECNOLOGICO DE CD ALTAMIRANO.

UNIDAD # 9 "TRANSFERENCIA DEL MATERIAL GENÉTICO".
LICENCIATURA EN BIOLOGIA QUINTO SEMESTRE.
QUE PRESENTA:
CANDIDO TORRES SANTIBAÑEZ.
CON # DE CONTROL: 09930058.
PROFESOR: FRANCISCO JAVIER PUCHE ACOSTA.
ASIGNATURA: BIOLOGIA MOLECULAR.
JUNIO/2012.
UNIDAD # 9"TRANSFERENCIA DEL MATERIAL GENÉTICO".
9.1 MECANISMOS DE TRANSFERENCIA NATURAL:
9.1.1 TRANSFORMACIÓN.
9.1.2 CONJUGACIÓN.
9.1.3 TRANSDUCCIÓN.
9.1.4 RECOMBINACIÓN.
9.1.5 TRANSFECCIÓN.
9.2 MECANISMOS DE TRANSFERENCIA ARTIFICIAL:
9.2.1 FÍSICOS
9.2.2 QUÍMICOS
OBJETIVO(S) GENERAL DEL CURSO
Entenderá las bases moleculares del control de la expresión génica y de la manipulación del ADN mediante el empleo de las diferentes técnicas moleculares que le permitan desarrollar proyectos de investigación básica y aplicada.
OJETIVO DE LA UNIDAD # 9.
Entender las bases moleculares del intercambio del material genético entre los diferentes seres vivos para su posterior aplicación
INTRODUCCIÓN
La transferencia de material genético entre microorganismos de sistemas acuáticos ha tenido un gran interés debido al surgimiento de bacterias multirresistentes a antibióticos.
El proceso de conjugación es considerado el principal camino para la transferencia horizontal de genes de resistencia entre bacterias.
La transferencia de plásmidos requiere el contacto directo entre célula y célula, lo cual resulta en una transferencia unidireccional del material genético, de la célula donadora a la receptora. Las bacterias que adquieren plásmidos R, pueden infectar al humano, directamente por contacto con el hospedero o a través de su ingesta por lo que el problema va más allá de la acuicultura y se convierte además en un
problema de Salud Pública.
Abstrac:
The transfer of genetic material among microorganisms of aquatics systems has had a great interest, due to the sprouting of bacteria multiresist to antibiotics. The process of conjugation is considered a major pathway for horizontal transfer of genes of resistance among bacteria.
The transfer of plasmids requires cell to cell contact resulting in unidirectional transfer of genetic material from a donor to a recipient cell. The bacteria that acquire R plasmid, they can infect human, directly by contact with the host or through its consumption for which the problem goes beyond the aquaculture and is converted besides in a problem
of Public Health.
METODOLOGIA.
La metodología que se utiliza son los pasos a seguir para la manifestación y desenlace de la unidad # 6 y con sus correspondientes subtemas: realizando enlaces de acuerdo a lo manifestado en clases de la unidad por el maestro y hacer conclusiones a lo entendido al final de la unidad de todos los subtemas, publicando lo más interesante yde cada tema visto en clase. y trabajos o ensayos obtenidos correspondiente a cada clase.
EVIDENCIAS.
LAS BACTERIAS
son microorganismos unicelulares que presentan un tamaño de unos pocos micrómetros (entre 0,5 y 5 μm, por lo general) y diversas formas incluyendo esferas (cocos), barras (bacilos) y hélices (espirilos).
Las bacterias son procariotas y, por lo tanto, a diferencia de las células eucariotas (de animales, plantas, hongos, etc.), no tienen el núcleo definido ni presentan, en general, orgánulos membranosos internos.
Generalmente poseen una pared celular compuesta de peptidoglicano. Muchas bacterias disponen de flagelos o de otros sistemas de desplazamiento y son móviles. Del estudio de las bacterias se encarga la bacteriología, una rama de la microbiología.
Las bacterias son los organismos más abundantes del planeta. Son ubicuas, se encuentran en todos los hábitats terrestres y acuáticos; crecen hasta en los más extremos como en los manantiales de aguas calientes y ácidas, en desechos radioactivos, en las profundidades tanto del mar como de la corteza terrestre. Algunas bacterias pueden incluso sobrevivir en las condiciones extremas del espacio exterior.
Se estima que se pueden encontrar en torno a 40 millones de células bacterianas en un gramo de tierra y un millón de células bacterianas en un mililitro de agua dulce. En total, se calcula que hay aproximadamente 5×1030 bacterias en el mundo.
Las bacterias son imprescindibles para el reciclaje de los elementos, pues muchos pasos importantes de los ciclos biogeoquímicos dependen de éstas. Como ejemplo cabe citar lafijación del nitrógeno atmosférico. Sin embargo, solamente la mitad de los filos conocidos de bacterias tienen especies que se pueden cultivar en el laboratorio, por lo que una gran parte (se supone que cerca del 90%) de las especies de bacterias existentes todavía no ha sido descrita.
En el cuerpo humano hay aproximadamente diez veces tantas células bacterianas como células humanas, con una gran cantidad de bacterias en la piel y en el tracto digestivo. Aunque el efecto protector del sistema inmunitario hace que la gran mayoría de estas bacterias sea inofensiva o beneficiosa, algunas bacterias patógenas pueden causar enfermedades infecciosas, incluyendo cólera, sífilis, lepra, tifus, difteria, escarlatina, etc.
Las enfermedades bacterianas mortales más comunes son las infecciones respiratorias, con una mortalidad sólo para la tuberculosis de cerca de dos millones de personas al año.
En todo el mundo se utilizan antibióticos para tratar las infecciones bacterianas. Los antibióticos son efectivos contra las bacterias ya que inhiben la formación de la pared celular o detienen otros procesos de su ciclo de vida. También se usan extensamente en la agricultura y la ganadería en ausencia de enfermedad, lo que ocasiona que se esté generalizando laresistencia de las bacterias a los antibióticos. En la industria, las bacterias son importantes en procesos tales como el tratamiento de aguas residuales, en la producción de queso, yogur,mantequilla, vinagre, etc., y en la fabricación de medicamentos y de otros productos químicos.
Aunque el término bacteria incluía tradicionalmente a todos los procariotas, actualmente la taxonomía y la nomenclatura científica los divide en dos grupos. Estos dominios evolutivos se denominan Bacteria y Archaea (arqueas).
La división se justifica en las grandes diferencias que presentan ambos grupos a nivel bioquímico y en aspectos estructurales.




Morfología y estructura.
Las bacterias son microorganismos procariotas de organización muy sencilla. La célula bacteriana consta:
Las bacterias son microorganismos procariotas de organización muy sencilla. La célula bacteriana consta:
citoplasma.
Presenta un aspecto viscoso, y en su zona central aparece un nucleoide que contiene la mayor parte del ADN bacteriano, y en algunas bacterias aparecen fragmentos circulares de ADN con información genética , dispersos por el citoplasma: son los plásmidos.
La membrana plasmática presenta invaginaciones, que son los mesosomas, donde se encuentran enzimas que intervienen en la síntesis de ATP, y los pigmentos fotosintéticos en el caso de bacterias fotosintéticas.
En el citoplasma se encuentran inclusiones de diversa naturaleza química.
Muchas bacterias pueden presentar flagelos generalmente rígidos, implantados en la membrana mediante un corpúsculo basal. Pueden poseer también, fimbrias o pili muy numerosos y cortos, que pueden servir como pelos sexuales para el paso de ADN de una célula a otra
Poseen ARN y ribosomas característicos, para la síntesis de proteínas.
pared celular es rígida y con moléculas exclusivas de bacterias.
REPRODUCCIÓN.
Tras la duplicación del ADN, que está dirigida por la ADN-polimerasa que se encuentra en los mesosomas, la pared bacteriana crece hasta formar un tabique transversal separador de las dos nuevas bacterias.
Pero además de este tipo de reproducción asexual, las bacterias poseen unos mecanismos de reproduccion sexual o parasexual, mediante los cuales se intercambian fragmentos de ADN
Generalmente las bacterias se reproducen por bipartición, como se ve en el siguiente esquema

9.1 MECANISMOS DE TRANFERENCIA NATURAL.
9.1.1 TRANSFORMACÓN.
En determinadas condiciones fragmentos de ADN exógeno o ADN transformante (tamaño superior a 3 x 105 dalton y longitud comprendida entre 5 x 106 y 15 x 106, que equivale a 200.000 pares de bases) con estructura helicoidal intacta pueden unirse a células bacterianas competentes y entrar en su interior. La entrada de estos segmentos necesita de la presencia de iones de k+, Mg++ y Ca++. El ADN entra en el espacio periplasmático, entre la pared celular y la membrana plasmática, allí una endonucleasas corta las dobles hélices en fragmentos de menor tamaño, posteriormente se degrada una de las dos hélices, de manera que lo que entra en el citiplasma es ADN de una hélice (monocatenario). Estos fragmentos de ADN monocatenario o ADN transformante pueden sustituir a fragmentos de ADN homólogo del cromosoma principal bacteriano mediante un mecanismo especial de recombinación. La recombinación genética tiene lugar entre el ADN transformante y el ADN de la bacteria receptora y se detecta por la aparición de bacterias descendientes transformadas para algún carácter. La existencia de este mecanismo permite construir Mapas genéticos de transformación.
9.1.2 CONJUGACIÓN.
En este proceso, una bacteria donadora F+ transmite a través de un puente o pili, un fragmento de ADN, a otra bacteria receptora F-. La bacteria que se llama F+ posee un plásmido, además del cromosoma bacteriano.
En la conjugación, el intercambio de material genético necesita de un contacto entre la bacteria dadora y la bacteria receptora. La cualidad de dador está unida a un factor de fertilidad (F) que puede ser perdido. La transferencia cromosómica se realiza generalmente con baja frecuencia. No obstante, en las poblaciones F+, existen mutantes capaces de transferir los genes cromosómicos a muy alta frecuencia.
La duración del contacto entre bacteria dadora y bacteria receptora condiciona la importancia del fragmento cromosómico transmitido. El estudio de la conjugación ha permitido establecer los mapas cromosómicos de ciertas bacterias. Ciertamente, la conjugación juega un papel en la aparición en las bacterias de resistencia a los antibióticos.



Tenemos una cepa receptora junto con material genético que aporta otra célula, de forma que a partir de ahí, puede producirse la conjugación, que es la transferencia de material; aunque para que sea viable, debe darse la recombinación gracias al proceso del entrecruzamiento. El intercambio genético no tiene lugar entre dos genomas completos como ocurre en eucariota, sino que, tiene lugar entre un genoma completo (F-) que se denomina endogenote, y otro incompleto, del donante, denominado exogenote. Obtendremos un merocigoto (diploide parcial). La genética bacteriana es la genética de la merocigosis.
Para que la recombinación dé algún cromosoma viable (que podrá ser circular), debe producirse un número par de entrecruzamientos, puesto que si no, no obtendremos ningún producto viable, sino un cromosoma largo lineal y extraño, parcialmente diploide. Si se da este número par de entrecruzamientos, obtendremos dos productos, uno que se perderá durante el crecimiento celular y otro que será viable.
Podemos encontrar procesos relacionados con la conjugación, tales como la sexducción, en el que se usan elementos F´ para crear diploides parciales. El F´ es un caso de factor F modificado, encontrándose en algunas cepas Hfr. Podemos obtener estirpes F´ con fragmentos muy grandes del cromosoma bacteriano. Los factores F´ son más fáciles de recombinar porque poseen más puntos para recombinar.
9.1.3 TRANSDUCCIÓN.
La transducción es la transferencia de genes de una bacteria a otra por medio de un virus. La incorporación de genes bacterianos al interior de la cápsida de un fago se produce a consecuencia de errores cometidos durante el ciclo duplicativo del virus. Cuando el virus que contiene estos genes infecta a una nueva bacteria, éste tienen la capacidad de transferirlos al cromosoma de ésta. La transducción es el mecanismo más frecuente de intercambio y recombinación genética en las bacterias. Hay dos tipos diferentes de transducción: la generalizada y la especializada.
Transducción generalizada
Ocurre durante el ciclo lítico (ciclo donde el fago rompe la bacteria) de los fagos y es capaz de transferir cualquier parte del genoma bacteriano. Durante la fase de ensamblaje viral, fragmentos del cromosoma bacteriano pueden quedar en la cápsida viral. Cuando este fago infecta a una nueva bacteria, el material puede ser transferido al cromosoma bacterial. La cantidad de DNA bacteriano trasportado depende principalmente del tamaño de la cápsida del virus.
Transducción especializada
En la transducción especializada, la partícula viral modificada transporta porciones específicas del genoma bacteriano. Los genes transportados son Bio y Gal. Este proceso es posible debido a un error durante el ciclo liso génico. Esto ocurre cuando se induce a un fago a abandonar el cromosoma de la célula huésped observándose en ocasiones una escisión incorrecta. Esto hace que el fago se lleve uno de los 2 genes localizados en sus extremos (Bio y Gal). El genoma viral resultante contiene porciones del cromosoma bacteriano justo al lado del sitio de la integración.
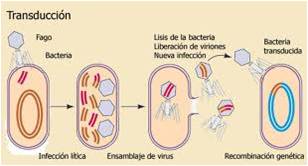
En otro sentido la Transduccion se define como la trasferencia de ADN de célula donadora a otra receptora mediatizado por partículas de bacteriófagos que contienen ADN genómico de la primera. En la transducción podemos distinguir dos estapas diferenciadas:
1. Formación de la partícula fágica transductora:
un trozo de material genético de la célula donadora se introduce en el interior de la cabeza de la cápsida de un fago. Las partículas transductoras son en cierta manera “subproductos” anómalos del ciclo normal del fago.
2. La partícula transductora inyecta de forma habitual el ADN que porta a la célula receptora, donde este ADN puede eventualmente recombinarse y expresar su información.
La transducción descubierta por Lederberg y Zinder se llama transducción generalizada.
Mediante ella se puede transferir cualquier marcador del genóforo del donador, con aproximadamente la misma frecuencia relativa (de ahí el calificativo de generalizada).
La transducción generalizada se produce sólo como consecuencia de infecciones líticas.
El ADN del genomio de la bacteria donadora que es introducido en la partícula transductora suele ir sin acompañamiento de ADN del propio fago.
Por ello, a esta peculiar partícula consistente en cápsida del fago que encierra sólo ADN genofórico de la bacteria se la denomina pseudovirión.
Siguiendo con la buena racha de descubrimientos, pocos años más tarde (1956), el mismo Lederberg (esta vez junto con su mujer, y con Morse) hallaron un tipo nuevo de transducción, mientras estaban estudiando el sistema del fago moderado l y su hospedador, E. coli. Este tipo de transducción recibió el nombre de transducción especializada, y sus caracteres distintivos son:
Sólo se transfieren marcadores cromosómicos cercanos al sitio de integración del ADN del fago (profago) en la célula lisogénica (p. ej., en el caso de l, los marcadores gal o bio).
Se produce únicamente como consecuencia de la inducción de la célula lisogénica por escisión del profago y consiguiente entrada a fase lítica, productora de nuevas partículas de fago.
El ADN genómico de la bacteria transportado por la partícula transductora va unido a ADN del fago.
La célula transductante se suele convertir en lisogénica para el fago correspondiente.



TRANSDUCCIÓN GENERALIZADA.
Se caracteriza porque en ella se puede transferir cualquier trozo de genóforo bacteriano, con tal de que tenga un tamaño compatible con la capacidad de “empaquetado” de ADN de la cápsida del fago. La partícula transductora (pseudovirión) se forma por empaquetamiento anómalo de ADN genofórico bacteriano. En el interior de la cabeza del pseudovirión sólo existe ADN bacteriano, sin ADN del fago. Las partículas transductoras sólo se forman como consecuencia de infecciones líticas del fago.
9.1.4 RECOMBINACIÓN.
Hasta ahora solo hemos estudiado el proceso de transferencia del material genético entre individuos que participan en un cruzamiento,. La existencia de esta transferencia se infiere a partir de la aparición de recombinantes entre los descendientes del cruzamiento. Sin embargo antes de que aparezca un recombinante estable, los genes trasferidos deben integrarse o incorporarse en el genomio de la célula recipiente mediante un mecanismo de intercambio.
Ahora veremos las características de este intercambio.
El intercambio en procariotas no ocurre entre dos genomios completos (como en eucariotas) sino si no entre un genomio completo llamado endogenote (el de la célula F-), y otro incompleto del donante llamado exogenote. De hecho lo que se obtiene en un diploide parcial o merocigoto.
Un solo hecho de recombinación no seria util para generar recombinantes viables, por que el circulo se rompería y formaria un cromosoma lineal extrañ, parcialmente diploide.
Para mantener intacto le circulo debe producirse par de cruzamientos. el fragmento producido se pierde durante el crecimiento celular. Por tanto no sobrevivevn los dos reciprocos de la reecombinacion, sino uno solo de ellos.
La recombinación en virus se descubrió por Delbrück, Bailey y Hershey en 1946. Este hallazgo fue posible a la existencia de variabilidad originada por mutación en caracteres de los virus fácilmente observables en los medios de cultivo. Dentro de este tipo de mutaciones están los caracteres de placa, el rango de hospedador, la sensibilidad a la temperatura, etc..).
Entre los caracteres de placa que se pueden analizar están la morfología de las calvas o halos de lisis que producen los virus al matar a las bacterias de cultivo en césped.

9.1.5 TRANSFECCIÓN.
Las técnicas de transfección celular, que se han desarrollado fundamentalmente para permitir la introducción de ácidos nucleicos en el interior de las células, han permitido en gran medida ampliar los conocimientos acerca de la regulación génica y de la función de las proteínas en los sistemas celulares. Actualmente se emplean en gran número de aproximaciones experimentales, en la generación de animales transgénicos, en la selección de líneas celulares modificadas, etc...
La introducción de una construcción de DNA recombinante en la que se ha situado el CDS (secuencia codificante) de un gen reportero (luciferasa, 'green fluorescent protein', beta-galactosidasa, cloramfenicol acetiltransferasa -CAT-, etc..) bajo una secuencia de regulación que se desea estudiar permite medir con facilidad tasas de expresión génica en diferentes situaciones experimentales.
Por el contrario, la introducción de un plásmido que contienen la secuencia codificante (CDS) de una proteína de interés bajo el control de un promotor (constitutivo, regulado, etc...) permite la producción de la proteína deseada que puede estar o no etiquetada ('tagged). En este caso se emplea la célula como una factoria de síntesis de proteínas a la que se le introduce en forma de plásmido la información de la proteína que se desea sintetice.
Tanto en el primero como en el segundo caso puede ser importante seleccionar las células que han adquirido el plásmido. Para facilitarlo se incluyen en éstos genes de resistencia a drogas que permiten a las células que los han adquirido sobrevivir en medios selectivos. Uno de los sistemas de selección más empleado es el de la resistencia a G418 (resistencia a neomicina) que permite seleccionar clones celulares de expresión estable. Así diferenciaremos entre la transfección temporal o transiente y la transfección estable o de larga duración.
Las técnicas de transfección actuales se pueden clasificar en los llamados métodos físicos (se basan en el uso de sistemas mecánicos, no biológicos, para lograr la inserción de material genético en las células) y los métodos químicos.
Las diferencias entre métodos químicos y físicos se encuentran a nivel metodológico, no a nivel de resultados, ya que el conjunto de métodos físicos no se basa en ningún sistema biológico, sino que pretende una inserción a “la fuerza”, de ese material genético. También hay que decir que este conjunto de técnicas sólo es funcional in vitro, ya que es necesaria la exposición de las células a medios extremos.
En otro punto de vista la transfección consiste en la introducción de material genético externo en células eucariotas mediante plásmidos, vectores víricos(en este caso también se habla de transducción) u otras herramientas para la transferencia.
El término transfección para métodos no virales se usa en referencia a células de mamífero, mientras que el término transformación se prefiere para describir las transferencias no virales de material genético en bacterias y células eucariotas no animales como hongos, algas o plantas.
La transfección de células animales generalmente se lleva a cabo abriendo poros o "agujeros" transitorios en la plasmática de las células mediante electroporación, para permitir el paso del material genético (como construcciones de DNA superenrollado o siRNA) aunque pueden ser transfectadas incluso proteínas (como anticuerpos, por ejemplo). Además de la electroporación, se pueden utilizar otras técnicas para efectuar la transfección, como por ejemplo liposomas producidos mediante la mezcla de lípidos catiónicos con el material genético, que se fusionarán con la membrana plasmática celular y depositarán su carga adentro.
MÉTODOS
Hay varios métodos para introducir DNA en una célula eucariota. Se han usado muchos materiales como vehículos para la transfección, divisibles en tres tipos: polímeros (catiónicos), liposomas y nano partículas.
Uno de los métodos más baratos (y fiables) es la transfección mediante fosfato de calcio, originalmente descubierta por F. L. Graham y A. J. van der Eb en 1973.Una solución salina tamponada con HEPES y que contiene iones fosfato se combina con una solución de cloruro de calcio que contiene el DNA a transfectar. Cuando ambas se combinan, se forma un precipitado fino formado por el calcio cargado positivamente y el fosfato cargado negativamente, que el DNA en su superficie.
Esta suspensión se añade a las células que se quieren transfectar (normalmente un cultivo celular en mono capa). Mediante un proceso no comprendido completamente, las células toman parte del precipitado, y junto con él, el DNA.
Otros métodos usan compuestos orgánicos altamente ramificados, los llamados dendrímeros, para unir el DNA e introducirlo en la célula. Un método muy eficiente es la inclusión del DNA en liposomas, pequeños cuerpos formados de una membrana en cierto modo similar a la membrana plasmática de la célula y que puede fusionarse con la misma, liberando el DNA al interior celular. Con células eucariotas, la transfección basada en la interacción lípido-catión es la más comúnmente utilizada, ya que son más sensibles a este método.
Otro método es el uso de polímeros catiónicos (o poli cationes) como DEAE-dextrano o polietilenimina. El DNA, cargado negativamente, se une al poli catión y el complejo es endocitado por la célula.
Una aproximación directa a la transfección es la balística o gene gun (pistola génica), en la que el DNA se une a una nano partícula compuesta de algún sólido inerte, generalmente oro, que es "disparada" directamente en el núcleo de la célula diana. El DNA puede también ser introducido en las células usando un virus como vector. En estos casos, la técnica es llamada transducción, y se dice que las células se transducen.
Otros métodos de transfección son la nucleofección, electroporación o magnetofección.

9.2 MECANISMOS DE TRANSFERENCIA ARTIFICIAL.
9.2.1 FISICOS
Los métodos físicos utilizan técnicas como la microinyección con una fina aguja de cristal y la electroporación (exposición de las células aun choque eléctrico).
La microinyección;
resulta ser una técnica muy potente y eficaz, ya que 1 de cada 5 células consigue incorporar el DNA foráneo de forma permanente. El problema de esta técnica es que exclusivamente se puede inyectar una célula cada vez, y por tanto no es el más recomendable para una terapia médica debido a que es demasiado laboriosa como para obtener un número de células indicadas (suficientes, aproximadamente un millar o más) para que el tratamiento tenga éxito.
Es una técnica muy efectiva aunque laboriosa. Es el método que se emplea en la introducción del DNA recombinante en las células embrionarias en el proceso de obtención de animales transgénicos.

La electroporación;
es una técnica que crea o que dota a la membrana de cierta permeabilidad al DNA durante unos pocos segundos debido a una descarga eléctrica. Si la célula o grupo de células sometidas, a dicho choque eléctrico están sumergidas en un medio rico en plásmidos, alguno de ellos puede penetrar en una célula. El problema que suscita este método es que los choques eléctricos no pueden producir los efectos deseados en algunas células y dañarlas o matarlas debido a esta causa.
Una vez introducida la molécula de DNA recombinante en el interior de las células producirá su efecto permitiendo, por ejemplo :
- Seleccionar células que hayan incorporado el DNA. Este sería el caso del aislamiento de clones celulares que presentaran expresión de un determinado producto. Es posible por la incorporación en el propio vector de marcadores de selección, típicamente la resistencia de G418 (neomicina),
- A las pocas horas o dias detectar la presencia de la proteína codificada por el plásmido.

9.2.2 QUIMICOS
Basados en la formación de complejos que las células sean capaces de adquirir e incorporar, bien sea directamente mediante la ruta endocítica (fosfato cálcico, DEAE dextrano) o a las membranas (lipofección).
Actualmente uno de los campos de estudio más importantes por lo que se refiere a les técnicas de transferencia de genes, son los estudios realizados con los llamados vectores sintéticos (también usados en técnicas de terapia génica), con tal de evitar los problemas derivados de la utilización de virus para la transferencia de genes.
Los vectores sintéticos (han tenido una alta eficacia in vitro, pero una baja in vivo) son de producción sencilla, altamente estables, y se pueden conseguir grandes construcciones.
Método del fosfato cálcico:
Basado en la obtención de un precipitado entre el cloruro de calcio y el DNA en una solución salina de fosfatos. En esta situación co-precipitan formando unos agregados que son endocitados/fagocitados por las células. Aparentemente el agregado con calcio protege al DNA de la degradación por las nucleasas celulares. El tamaño y la calidad del precipitado es crítico para el éxito del proceso, que se ve afectado por factores tales como pequeños cambios en el pH de la solución, etc...
Método del DEAE dextrano:
Basado en la obtención de complejos entre la resina DEAE y el DNA. Los polímeros de DEAE dextrano o polybreno tienen una carga que les permite unirse a las muy negativamente cargadas moléculas de DNA. El DNA acomplejado se introduce en las células mediante choque osmótico mediante DMSO o glicerol. El uso de DEAE dextrano se limita a las transfecciones transientes.
Método DNA desnudo:
El DNA desnudo (técnica en fase altamente experiemtal) es incapaz de entrar en una célula y aún consiguiendo entrar en ellas es rápidamente degradado. La línea de investigación busca incorporar esas secuencias de DNA a un transportador, que le encierra y le lleva hasta la célula Diana en dónde a través de interacciones de membrana se asocia con ella para entregar el material al interior.
Los problemas que encontramos son que el DNA (rico en cargas -) por ser estructura polar tiene muchas dificultades para cruzar la membrana plasmática. Además el DNA codificante para 1 gen es aproximadamente de 10 kb, esto representa la longitud aproximada de una célula, en el caso de encontrar el DNA sin ningun tipo de compactatción.
Los problemas que encontramos son que el DNA (rico en cargas -) por ser estructura polar tiene muchas dificultades para cruzar la membrana plasmática. Además el DNA codificante para 1 gen es aproximadamente de 10 kb, esto representa la longitud aproximada de una célula, en el caso de encontrar el DNA sin ningun tipo de compactatción.
Método Péptidos fusiogénicos:
Los péptidos fusiogénicos surgen en una linea de trabajo que nos permita paliar aquellas características del DNA desnudo que nos impiden la entrada. Estos péptidos fusiogénicosa se utilizan para compactar DNA. y de este modo adoptan DNA con proteínas ricas en cargas +, del tipos Histonas.
Esto supone una condensación considerable, (visible con MET) ya que el DNA aparece en la forma de partículas de 50-150 nanometros, pero sólo un cierto grado de condensación permite que 1 transportador incluya 1 molécula de DNA. Asimismo la reducción de tamaño del DNA hasta unos 20 nanometros facilitaría el paso a la célula y hasta el núcleo, incrementando su protección contra las nucleases citoplasmáticas. En el proceso de penetración celular, los transportadores sintéticos (si su carga neta es +) interactuan con Aminoazucares de la Membrana plasmáticacon (carga -) para producir la incorporación a la célula en forma de endosomas. Por tanto tenemos un problema más a superar.
También se ha visto que los puntos en dónde se da una mayor transferencia son los pulmones y el hígado, con lo que se considera que los transportadores se agrupan en grandes complejos y que interactuan con proteínas de la sangre, con lo que no abandonan eficazmente el sistema vascular, finalmente son retenidos por los filtros naturales del cuerpo, tales como los pulmones o el hígado, en dónde interactuan. Esto marca dos puntos a superar en este modelo de transferencia de genes in vivo, en primer lugar el superar los filtros naturales que presenta todo organismo, en segundo punto dirigir el transportador dentro del cuerpo con tal de incrementar la afinidad a las células diana.
Una solución propuesta a este último problema es unirlos a ligandos específicos, tales como azúcares, aminoacidos, hormonas,.... Una vez en el tejido adecuado, el proceso de absorción se da por endocitosis formandose un endosoma (en este punto el transportador con el vector han de salir al citoplasma, ya que si no recibirán ataques lisosomales que digerirán el conjunto). Para conseguir una salida se acopla al vector un péptido fusiogena (p.ej. una molécula DOPE, que permite la salida del endosoma por desestabilitzación de les Mbs).
CONCLUSIÓN.
Exelente profesor.....francisco javier puche acosta.
BIBLIOGRAFIA:
BIBLIOGRAFIA:
No hay comentarios:
Publicar un comentario